
 D.P.Barlow & M.S.Bartolomei @ CSH
D.P.Barlow & M.S.Bartolomei @ CSH

Öz
Genomik damgalama, memelilerde genlerin bir altkümesini etkiler ve tek-allelli (İng. monoallelic), ebeveyne-özgü bir ifade örüntüsüyle sonuçlanır. Bu genlerin çoğu, yalıtıcıların veya uzun kodlamayan RNA'ların (İng. long noncoding RNA - lncRNA) kullanımı yoluyla düzenlenen (regüle edilen) kümelerde bulunur. Ebeveynsel allelleri ayırt etmek için damgalanmış genler, DNA metilasyonu kullanımı yoluyla gametlerdeki damgalama kontrol elemanlarında epigenetik olarak işaretlenir. Damgalanmış gen ifadesi, daha sonra lncRNA'lar, histon modifikasyonları, yalıtıcılar ve daha yüksek mertebeden kromatin yapısı yoluyla sunulur. Memeli genomunun geniş çaplı yeni baştan programlanmasına rağmen, böyle damgalamalar, bu mekanizmalar yoluyla döllenmeden sonra korunur. Genomik damgalama, memeli epigenetik düzenlemesinin (regülasyonunun) kavranması için harika bir modeldir.
Genel Bakış
Memeliler diploid organizmalardır; hücrelerinde iki uyumlu kromozom kümesi bulunur: Biri anneden, biri de babadan kalıtılır. Dolayısıyla memelilerde her genden iki kopya vardır. Normalde her bir genin hem annesel hem de babasal kopyasının, herhangi bir hücrede etkin olma potansiyeli aynıdır. Genomik damgalama, bu potansiyeli değiştiren bir epigenetik mekanizmadır; çünkü iki ebeveynsel kromozomdan birindeki genin ifadesini kısıtlar. Bu, genomumuzdaki yaklaşık 25.000 tane genin sadece birkaç yüz tanesi tarafından sergilenen bir görüngüdür; çoğunluk iki ebeveynden de kalıtıldığında eşitçe ifade edilir. Genomik damgalama, hem erkek hem de dişi yavruları etkiler ve dolayısıyla ebeveyensel kalıtımın bir sonucudur; cinsiyetin değil. Bunun anlamını bir örnekle açıklayalım: Anneden kalıtılan bir kromozom üzerinde etkin olan bir damgalanmış gen, erkek yavruların da dişi yavruların da hepsinde annesel kromozomda etkin olup, babasal kromozomda suskun olacaktır.
Genomik damgalamanın tanımı, burada "diploid hücrelerde ebeveyne-özgü gen ifadesi" ile kısıtlanmıştır. Dolayısıyla, tüm genlerden iki ebeveynsel kopya içeren diploid hüceler, damgalanmış bir genin sadece bir ebeveynsel kopyasını ifade edip, diğer ebeveynsel kopyasını susturacaktır. Tersine, damgalanmamış genler diploid bir hücrede iki ebeveynsel gen kopyasından da ifade edilir. Genomik damgalama kavramını anlamak için damgalanmış genler ile embriyoya eşitsiz ebeveynsel genetik katkı nedeniyle belirgin ebeveyne-özgü ifade sergileyen genleri ayırt etmek önemlidir. Eşitsiz ebeveynsel genetik katkıya örnek olarak şunlar verilebilir: Sadece erkeklerde bulunan Y-kromozomu bağlantılı genler, dişilerde X inaktivasyonundan kaçan genler (X-bağlantılı gen ürünlerinden, erkeklere kıyasla iki kat fazla üretilmesine neden olur), temelde anne ebeveynden gelen mitokondriyal genler ve sadece sperm veya yumurta sitoplazmasında bulunan haberci RNA'lar (mRNA'lar) ile proteinler.
Memelilerdeki genomik damgalamanın çoğu özelliği, onu post-genomik dönemlerde çok ilginç bir biyolojik problem yapar. Genomik damgalamaya konu olan gen altkümesinin, büyük ölçüde, embriyonik ve yenidoğansal (İng. neonatal) büyümeyi düzenleyen faktörleri kodlaması merak uyandırıcıdır. Dolayısıyla, genomik damgalamanın, memeli üremesinde belli bir rolü oynamak üzere evrilmiş olması muhtemeldir. Ayrıca, annesel ebeveynin bir içsel üreme sistemine uyumlanmasına (adaptasyonuna) olası bir evrimsel yanıt olarak, ebeveynsel çekişme konusunda ipucu sağlar. Yanı sıra, memeli genomunun kendisini işgalci DNA dizilimlerine karşı koruma şekline kısa bir bakış sağlayabilir. Genomik damgalama, entelektüel açıdan zorlayıcı bir görüngüdür; özellikle de diploid bir organizmanın, diploid durumun avantajlarından vazgeçen bir susturma sistemini neden geliştirdiği sorusunu doğurduğu için.
Bilgimizin bu aşamasında, genomik damgalama, dört ökaryotik âlem (Protistler, Mantargiller, Bitkiler, Hayvanlar) arasında yaygın gibi görünmemektedir. Bununla birlikte, iki omurgasız eklembacaklıda (Coccidae ve Sciaridae) ve mısır ile Arabidopsis gibi bazı tohumlu bitkilerin endospermlerinde, muhtemelen ilişkili bir biçimde, mevcuttur. Bu dağılım, canlılığın evrimi sırasında, genomik damgalamanın en azından üç kez bağımsız olarak ortaya çıktığına işaret eder. Şaşırtıcı şekilde, genomik damgalamanın bu öngörülen bağımsız evrimine rağmen, damgalama mekanizması arasında bazı benzerlikler görünür. Bunun, hem genomik damgalamanın hem de normal gen düzenlemenin altında yatan temel epigenetik düzenleme mekanizmalarının korunumunu yansıtması muhtemeldir.
1. Tarihsel Bakış
Memelilerde genomik damgalama olmasının, (1) genetik özelliklerin ve hastalıkların klinik yönetimi, (2) yardımcı üreme teknolojileriyle insan ve hayvan çoğalmasının kontol kapasitesi, (3) biyoteknolojinin ve post-genomik tıbbi araştırmanın ilerlemesi açılarından dikkate değer tıbbi, toplumsal ve entelektüel sonuçları vardır. İster araştırma ister tıp alanında olsun, günümüzde genetik problemler hakkındaki herhangi bir tartışmanın, bir genin iki-ebeveynsel (yani diploid) bir ifade modu mu sergilediğini, yoksa genomik damgalamayla ebeveyne-özgü (yani haploid) ifade mi sergilediğini hesaba katması gerekir. Genomik damgalamanın insan sağlığı ve esenliği için önemine rağmen, varlığının ve dikkate değerliğinin geniş çapta kabûlünün, farelerdeki üç genin ebeveyne-özgü ifade sergilediğinin su götürmez biçimde gösterildiği doksanlı yılların başına kadar gerçekleşmemiş olması şaşırtıcıdır.
Kromozomların bütün hâlinde ebeveyne-özgü davranışı, 1930'lar gibi erken bir tarihte, eklembacaklılardaki kromozomların sitogenetik incelenmesinde gözlemlenmiştir (Chandra & Nanjundiah 1990). İlginçtir; "kromozom damgalama" terimi ilk olarak, bazı eklembacaklı türlerde cinsiyet belirleniminde rol oynayan ebeveyne-özgü kromozom elemesini tanımlamak için ortaya atılmıştır (Crouse et al. 1971). Dişi keselilerin tüm hücrelerinde ve farenin ekstra-embriyonik dokularında, iki X-kromozomundan birinin ebeveyne-özgü inaktivasyonuna yol açan, memelilerdeki X-kromozomunun kromozomal damgalaması da fark edilmiştir (Cooper et al. 1971). Aynı dönemde, klasik genetikçiler, damgalanmış gen ifadesinin gözlemi için temel teşkil eden kromozomal translokasyonlar taşıyan mutant fareler üretiyordu. Başlangıçta kromozomlar üzerinde genlerin konumlarını haritalandırmak için kullanılan bu "translokasyon" farelerinden bazıları, bir ebeveynsel kromozomun yokluğunda (tek-ebeveynsel disomi olarak bilinir; İng. uniparental disomy - UPD; Fig. 1), belirli kromozomal bölgeler diğer ebeveynsel kromozomun ikilenmişleri olarak kalıtıldıklarında, ebeveyne-özgü bir fenotip sergiledi. Bu sonuçlar, "belirli annesel veya babasal genlerin haploid ifadesinin, normal fare gelişimi için önemli olması" olasılığına işaret etti (Searle & Beechey 1978). Aynı sıralarda, başka genetikçiler de kromozom-17'nin büyük bir delesyonunu (büyük bölümünün silinmesini) taşıyan ve "firkete kuyruklu" fare olarak bilinen sıradışı bir mutant fare kullanarak, "belli bir lokusta heterozigot olan organizmaların, hangi gametin genotipe hangi alleli kattığına bakmaksızın, fenotipik olarak özdeş oldukları" şeklindeki, genetiğin temel inançlarından birini su götürmez biçimde ortadan kaldırdı (Johnson 1974). Firkete kuyruklu delesyonunu annesel ebeveynden alan yavrunun büyüklüğü artıyor ve embriyonik gelişimin ortalarına doğru ölüyordu; genetik olarak özdeş kromozomun babasal aktarımı ise yaşayabilen ve üreyebilen fareler oluşturuyordu (Fig. 1). Daha önceden memelilerdeki damgalanmış X-kromozomu inaktivasyonunun tanımı yayımlanmış olmasına karşın, bu genetik translokasyon ve delesyon deneylerinin yeğlenen yorumunun, ilgili bölgelerin damgalanmış genler içerdiği değil, bu otozomlardaki genlerin birincil olarak haploid yumurta veya spermde devreye girip embriyonik gelişimde sonradan kullanılan proteinleri modifiye ettiği şeklinde olması dikkate değerdir. Buna rağmen, annesel ve babasal genomun işlevlerinin farklılık göstermesi kavramı yer ediniyordu ve "firkete kuyruklu kromozomal bölgede annesel genom normalde aktifken, babasal karşılığının tercihan inaktifleştirilmiş olabileceği" fikri öne sürüldü (McLaren 1979).
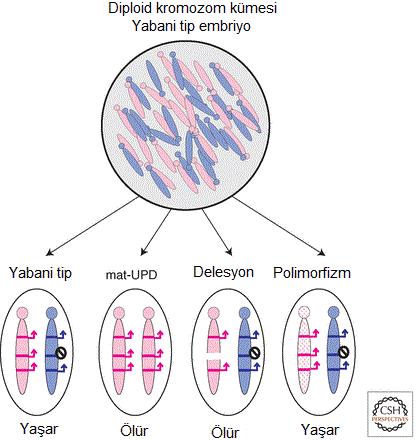 Figür-1. Genomik damgalamayı incelemk için annesel ve babasal kromozomların ayırt edilmesine olanak tanıyan fare modelleri. Memeliler diploid olup, annesel ve babasal ebeveynlerden tam birer kromozom kümesi kalıtır. Ancak, (1) ebeveynlerden birinden bir kromozom ikilisinin iki kopyasını da alıp, diğer ebeveynden hiç kopyasını almayan (UPD olarak bilinir) fareler, (2) ebeveynlerden birinden bir kısmi kromozomal delesyon alıp, diğer ebeveynden yabani tip kromozom alan fareler, (3) ebeveynlerden birinden tekil-nükleotid polimorfizmleri taşıyan kromozomlar alıp, diğer ebeveynden yabani tip kromozom alan fareler üretilebilir. UPD'li ve delesyonlu yavruların ölüme sebebiyet veren fenotipler sergilerken, SNP'lerin yaşayabilen yavrular üretmesi muhtemeldir.
Figür-1. Genomik damgalamayı incelemk için annesel ve babasal kromozomların ayırt edilmesine olanak tanıyan fare modelleri. Memeliler diploid olup, annesel ve babasal ebeveynlerden tam birer kromozom kümesi kalıtır. Ancak, (1) ebeveynlerden birinden bir kromozom ikilisinin iki kopyasını da alıp, diğer ebeveynden hiç kopyasını almayan (UPD olarak bilinir) fareler, (2) ebeveynlerden birinden bir kısmi kromozomal delesyon alıp, diğer ebeveynden yabani tip kromozom alan fareler, (3) ebeveynlerden birinden tekil-nükleotid polimorfizmleri taşıyan kromozomlar alıp, diğer ebeveynden yabani tip kromozom alan fareler üretilebilir. UPD'li ve delesyonlu yavruların ölüme sebebiyet veren fenotipler sergilerken, SNP'lerin yaşayabilen yavrular üretmesi muhtemeldir.Memelilerde genomik damgalamanın varlığının ortaya konması yolunda başlıca adımlardan biri, sadece fare yumurta hücresi çekirdeğinden diploid tek-ebeveynsel embriyoların üretilmesi olasılığını sınamak için kullanılan ileri bir çekirdeksel aktarım teknolojisinin geliştirilmesiyle birkaç yıl sonra geldi. Çekirdeksel aktarım tekniği, yeni döllenmiş bir yumurtadan bir donör erkek veya dişi pronükleus aldı ve ince bir mikro-pipet kullanarak onu konak bir döllenmiş yumurtanın içine yerleştirdi; konaktan ya annesel ya da babasal pronükleus çıkartılmıştı. Böylece yeniden diploid embriyolar üretildi ama bunların ya iki annesel genomu ya da iki babasal genomu vardı (bunlara sırasıyla jinogenetik ve androgenetik embriyo adları verildi; Fig. 2). Teknik ilk olarak, döllenmiş firkete kuyruklu mutant embriyolardan alınan çekirdeklerin, yabanıl tip bir konak yumurtaya aktarıldıklarında kurtarılamayacaklarını göstermek için kullanıldı. Bu, oosit sitoplazmasının değil, embriyonik genomun firkete kuyruk kusurunu taşıdığına ilişkin kanıt sağladı. Ayrıca, kromozom-17'nin annesel ve babasal kopyası üzerindeki genlerin, embriyonik gelişim sırasında farklı işlevler sergilediği iddiasını doğruladı (McGrath & Solter 1984b). Daha sonra çekirdeksel aktarım, iki annesel pronükleustan (jinogenetik) veya iki babasal pronükleustan (androgenetik) yapılandırılan embriyoların hayatta kalmayı başaramayıp, sadece bir annesel pronükleus ile bir babasal pronükleustan üretilen embriyoların yaşayabilir ve üreyebilir yavru verebildiğini göstermek için kullanıldı (McGrath & Solter 1984a; Surani et al. 1984). Bu çalışma, tek-ebeveynsel farelerin erişkinliğe ulaşabileceği şeklindeki eski iddiayı alabora etti (Hoppe & Illmensee 1982). Jinogenetik embriyoların ölümü sırasında döleşine (plasenta/ etene) katkı yapan ekstra-embriyonik dokuları kusurlu oluyordu; androgenetik embriyoların ise embriyonik dokuları kusurlu oluyordu. Bu sonuçlar şu hipoteze yol açtı: Embriyonik gelişim annesel genomdan ifade edilen damgalanmış genlere gereksinim duyarken, ekstra-embriyonik gelişim için gereken damgalanmış genler de babasal genomdan ifade edilir (Barton et al. 1984). Faredeki damgalanmış genlerin sonradan tanımlanması, damgalanmış genlerin işlevinde bir taraflılık (İng. bias) olduğunu doğrulamadı ama jinogenetik ve androgenetik embriyolar arasında gözlemlenen farkların, damgalanmış genlerden birinin ya da birkaçının baskın etkisi tarafından açıklanabileceğine işaret etti.
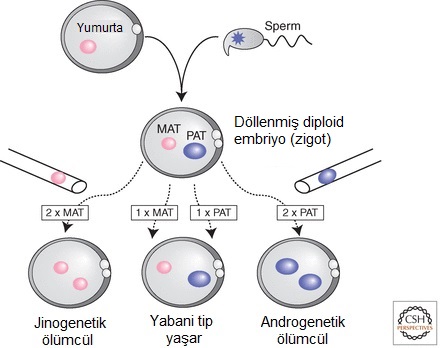 Figür-2. Memeli üremesi için bir annesel ve bir babasal genom gerekir. Çekirdeksel aktarım tekniği, mikropipetler ve yüksek güçlü mikroskoplar kullanılarak, yeni döllenmiş bir yumurtadan eril veya dişil çekirdeğin çıkarılmasını sağlar ve çekirdeği çıkarılmış ikinci bir "konak" döllenmiş yumurtaya onları çeşitli kombinasyonlarda yerleştimeyi sağlar. Böylelikle iki annesel (jinogenetik) veya iki babasal (androgenetik) genomları olan veya iki ebeveynli genomları (yabani tip) olan yeni diploid embriyolar üretilebilir. Erken embriyonik evrelerde, jinogenetik ve androgenetik embriyolar ölümcüldür. Sadece bir annesel ve bir babasal çekirden (yabani tip) alan yeni baştan oluşturulmuş embriyolar sağ kalıp yavru üretir. Bu deneyler, memeli üremesinde hem annesel hem de babasal genomun gerekliliğini gösterir ve iki ebeveynsel genomun farklı gen kümelerini ifade etmesinin, eksiksiz embriyonik gelişim için gerekliliğine işaret eder.
Figür-2. Memeli üremesi için bir annesel ve bir babasal genom gerekir. Çekirdeksel aktarım tekniği, mikropipetler ve yüksek güçlü mikroskoplar kullanılarak, yeni döllenmiş bir yumurtadan eril veya dişil çekirdeğin çıkarılmasını sağlar ve çekirdeği çıkarılmış ikinci bir "konak" döllenmiş yumurtaya onları çeşitli kombinasyonlarda yerleştimeyi sağlar. Böylelikle iki annesel (jinogenetik) veya iki babasal (androgenetik) genomları olan veya iki ebeveynli genomları (yabani tip) olan yeni diploid embriyolar üretilebilir. Erken embriyonik evrelerde, jinogenetik ve androgenetik embriyolar ölümcüldür. Sadece bir annesel ve bir babasal çekirden (yabani tip) alan yeni baştan oluşturulmuş embriyolar sağ kalıp yavru üretir. Bu deneyler, memeli üremesinde hem annesel hem de babasal genomun gerekliliğini gösterir ve iki ebeveynsel genomun farklı gen kümelerini ifade etmesinin, eksiksiz embriyonik gelişim için gerekliliğine işaret eder.Fare genetiğinden çıkan destekleyici verilerle bir araya getirilen çekirdeksel aktarım deneyleri, farelerde embriyojenez (embriyo oluşumu) için her iki ebeveynsel genoma da gerek olduğuna ilişkin ikna edici kanıt sağlayarak, memelilerde genomik damgalamanın varlığı için güçlü bir temel attı (Fig. 2). UPD (tek-ebeveynsel disomi) kromozomlar yaratmak için "translokasyon" farelerinin kullanıldığı (Fig. 1), embriyonik gelişime ebeveynsel kromozom katkısının geniş çaplı bir incelemesi, farelerdeki kromozom-2 ve kromozom-11 üzerinde, iki annesel ya da iki babasal kopya hâlinde bulunduğunda karşıt fenotipler gösteren iki bölge belirledi. Bu, memelilerde ebeveyne-özgü gen ifadesi savını daha da güçlendirdi (Cattanach & Kirk 1985). Buna ek olarak, insan verileri, başta Prader-Willi sendromu (özellikle babasal aktarımdan doğuyor görünür) olmak üzere bazı genetik durumların, en iyi açıklamasının ebeveyne-özgü gen ifadesi olduğuna işaret etti (Reik 1989). Döllenmiş bir fare yumurtasına gen dizilimlerinin mikro-enjeksiyonu ile transjenik fareler yapmak için yeni geliştirilen teknolojinin uygulandığı deneylerden başka ipuçları da geldi. Bu sıklıkla, somatik dokularda transgenin susturulmasını beklenmedik şekilde tetikleyen DNA metilasyonu problemiyle kuşatılıyordu. DNA metilasyonu edinme becerilerinde ebeveyne-özgü farklılıklar gösteren bazı transgenlerin bile olması, ebeveynsel kromozomların farklı davrandığı iddiasını artırıyordu. Bu normalde şu örüntüye uyuyordu: Annesel aktarımlı transgenler metillenirken, babasal aktarımlı transgenler metillenmiyordu. Bununla birlikte, sadece birkaç vakada DNA metilasyon farkları, ebeveyne-özgü ifade ile bağlaşıktı. "Transgen" metilasyon damgalaması ile endojen fare genlerinin genomik damgalaması arasında sonradan çok sayıda benzerlik bulunmasına karşın, çeşitli özellikler onları ayırır (Reik et al. 1990). Bunlar arasında, ipliğe-özgü arka plan etkilerine yüksek bir hassasiyet, farklı kromozomal integrasyon alanlarında damgalanmış ifadenin sürdürülememesi ve damgalama etkisi üretmek için yabancı DNA dizilimlerine gereksinim bulunur (Chaillet et al. 1995).
Destekleyici verilerin zenginliğine rağmen, memelilerde genomik damgalamanın varlığının nihai kanıtı, damgalanmış ebeveyne-özgü ifade sergileyen genlerin belirlenmesine bağlıydı. Bu 1991'de gerçekleşti; üç damgalanmış fare geni tanımlandı. Bunlardan ilki olan Igf2r (insülin-benzeri büyüme faktörü tip 2 almacı; büyüme hormonu insülin-benzeri büyüme faktörü tip2 için "temizleyici" bir almaç) annesel ifadeli bir damgalanmış gen olarak kimliklendirildi. Bu genin, firkete kuyruklu mutant farenin aşırı irileşen fenotipini açıkladığı daha sonra gösterildi (Barlow et al. 1991). Birkaç ay sonra, Igf2 geni babasal ifadeli bir damgalanmış gen olarak kimliklendirildi (DeChiara et al. 1991; Ferguson-Smith et al. 1991). Son olarak, H19 geni (cDNA klon numarası 19; bir fetal hepatik kütüphaneden yalıtılmış), alışılmadık bir uzun kodlamayan RNA (lncRNA) sonradan annesel ifadeli bir damgalanmış gen olarak tanımlandı (Bartolomei et al. 1991). Bu üç damgalanmış geni tanımlamak için, her biri fare genetiğinde yeni çıkan teknolojilere bağlı olan çeşitli stratejiler kullanıldı. Igf2r için, kromozom-17 üzerindeki firkete kuyruk delesyonuna haritalandırılan genleri tanımlamak için konumsal klonlama kullanıldı. Bunun ardından bir ebeveynden delesyonu kalıtım yoluyla alan fareler, anneye-özgü ifade sergileyen genleri belirlemek için kullanıldı (Fig. 1). Igf2 için, bu büyüme faktörünün embriyonik gelişimdeki fizyolojik rolü, gen nakavtı (İng. gene knockout) teknolojisiyle sınandı. Şaşırtıcı bir şekilde, mutant işlevsiz allel taşıyan fareler, babasal aktarıma uyan bir fenotip sergiledi ama annesel aktarımlı fenotip yoktu. H19 lncRNA geni, kromozom-7 üzerinde Igf2 lokusuna yakın olarak haritalandırıldıktan sonra bir damgalanmış gen olarak tanımlanarak, damgalanmış genlerin birlikte kümelenmiş olabilecekleri hipotezini kanıtladı. Damgalanmış genleri tanımlamak için daha sonraki girişimlerde bu stratejilerin yararlı olduğu kanıtlanmakla beraber, memelilerde genomik damgalamayı kontrol eden mekanizmanın anlaşılmasında en önemli keşif, damgalanmış genlerin sıkıca kümelenmiş olduklarının gösterilmesi oldu.
2. Genomik Damgalama: Bir Epigenetik Gen Düzenleme Sistemi
Genomik damgalamanın tanımlayıcı özelliği, cis-etkir olmasıdır (Latince "cis-" öneki "yanında, aynı yanda" anlamında gelir; "trans-" önekinin karşıtı olarak da kullanılır; "cis-etkir" sıfatı, yanındaki genlere yani aynı kromozomdaki genlere etkiyen genler için kullanılır). Dolayısıyla, damgalama mekanizması sadece bir kromozom üzerine etkir. Eğer popülasyon yakın akraba ebeveynlerden türemediyse, iki ebeveynsel kromozom normalde çok sayıda tekil baz çifti farklılığı (tekil-nükleotid polimorfizmleri ) içerir; ama eğer yakın akraba fare soyları kullanılırsa, genetik olarak özdeş olabilirler. Genomik damgalama, genetik açıdan özdeş ebeveynsel kromozomlara sahip olan yakın akrabalardan olma farelerde gerçekleştiğinden, DNA diziliminin taşıdığı bilgiyi modifiye edip iki ebeveynsel gen kopyası arasında ifade farklılığı yaratmak için sürecin bir epigenetik mekanizma kullanması gerektiği sonucuna varıldı. Bu gözlemler ayrıca cis-etkir, tek kromozomla kısıtlı bir susturma mekanizmasının işlemekte olduğuna ve böylece susturma faktörlerinin aktif gen kopyasına erişmek için çekirdekten rahatça geçemediğine işaret ediyordu. Her ne kadar damgalanmış genler bir ebeveynsel kromozomda diğerine göre baskılanmış olsa da, genomik damgalamanın illâ bir susturma mekanizması olması gerekmez ve ifadede ebeveyne-özgü farklar tetiklemek için gen düzenlemenin herhangi bir düzeyinde (yani başlatıcıda, yükselticilerde, birleştirme eklemlerinde veya poliadenilasyon alanlarında) işleme potansiyeli vardır.
 Memelilerde genomik damgalamanın anahtar özellikleri.
Memelilerde genomik damgalamanın anahtar özellikleri.O hâlde genomik damgalama, iki ebeveynsel kromozomdan birini modifiye eden yani "damgalayan" epigenetik sisteme bağlı olmalıdır (Fig. 3). Bu damga daha sonra yazımsal faktörleri veya mRNA işleme faktörlerini çekmek ya da itmek için kullanılır ve böylece bir ebeveynsel kromozomdaki damgalanmış genin ifadesini değiştirir. Soy içi çiftleşme sonucu doğmuş olduklarından genetik açıdan özdeş kromozomlara sahip olan fareler de genomik damgalama sergilediğinden, ebeveynsel damgaların embriyo diploid hâle geldikten sonra edinilmesi olası değildir; çünkü hücrelerin epigenetik mekanizmasının özdeş ebeveynsel gen kopyaları arasında ayrım yapmasının yolu olmaz. Dolayısıyla, ebeveynsel damgaların, iki ebeveynsel kromozom kümesi ayrıyken edinilmiş olması gerekir ve bu ancak gamet oluşumu sırasında ve döllenmeden sonraki ∼12 saat içinde gerçekleşir (Fig. 3). En olası senaryo, gametik damgaların babasal damgalı genlere sperm oluşumu sırasında ve annesel damgalı genlere yumurta oluşumu sırasında yerleştirilmesidir. "Damgalanmış" DNA diziliminin anahtar özelliklerinden biri şudur: İki ebeveynsel gametten sadece birinde modifiye edilir; dolayısıyla her biri farklı bir DNA dizilimine doğru yönelmiş olan, biri sperme-özgü diğeri oosite-özgü olmak üzere iki çeşit tanıma sistemi gerekir. Başka bazı özellikler de damganın gereksinimidir. Birincisi, oluşturulduğunda, döllenmeden sonra embriyo diploid olduğunda aynı ebeveynsel kromozom üzerinde kalmalıdır. İkincisi, damga, yetişkin hayvan ve embriyonun mitozu yoluyla sabit şekilde kalıtılmalıdır. Son olarak, silinebilir olmalıdır. Bu sonuncusu gereklidir çünkü embriyo ya eril ya da dişil bir gelişimsel yol izleyecektir ve gonadlarının sadece bir çeşit damgalanmış haploid ebeveynsel gamet üretmesi gerekir. O nedenle, embriyonik diploid hücrelerden doğan germ hücrelerinin (Fig. 3) önce kalıtımla aldıkları annesel ve babasal damgaları yitirmeleri, sonra gametinkini kazanmaları gerekir.
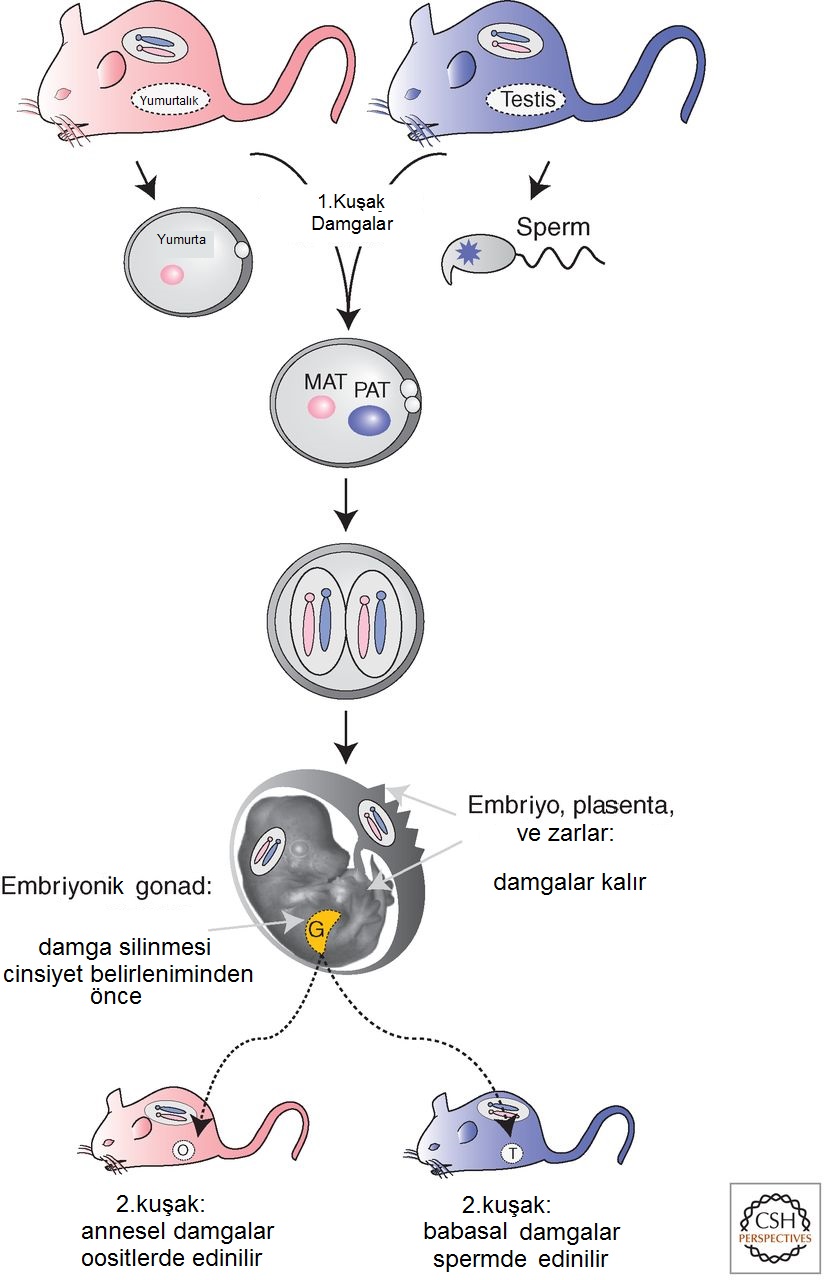 Figür-3. Memeli gelişiminde damga edinimi ve silinimi. Damgalar, gametlerce edinilir; dolayısıyla, oositler ve spermler damgalanmış kromozomlar taşır (ilk-kuşak damgalar). Döllenmeden sonra embriyo diploid iken, embriyonun, yumurta kesesinin, plasentanın ve yetişkinin her bir hücresinin bölünmesinden sonra damga aynı ebeveynsel kromozom üzerinde korunur. Germ hücreleri embriyonik gonadda oluşturulur ve damgalar sadece bu hücrelerde zinsiyet belirleniminden önce silinir. Embriyo erkekse gonadlar testise dönüşür ve kromozomları üzerinde babasal bir damga taşıyan haploid sperm üretir. Benzer biçimde embriyo dişiyse, yumurtalıklardaki kromozomlar annesel damgalar edinir (ikinci-kuşak damgalar).
Figür-3. Memeli gelişiminde damga edinimi ve silinimi. Damgalar, gametlerce edinilir; dolayısıyla, oositler ve spermler damgalanmış kromozomlar taşır (ilk-kuşak damgalar). Döllenmeden sonra embriyo diploid iken, embriyonun, yumurta kesesinin, plasentanın ve yetişkinin her bir hücresinin bölünmesinden sonra damga aynı ebeveynsel kromozom üzerinde korunur. Germ hücreleri embriyonik gonadda oluşturulur ve damgalar sadece bu hücrelerde zinsiyet belirleniminden önce silinir. Embriyo erkekse gonadlar testise dönüşür ve kromozomları üzerinde babasal bir damga taşıyan haploid sperm üretir. Benzer biçimde embriyo dişiyse, yumurtalıklardaki kromozomlar annesel damgalar edinir (ikinci-kuşak damgalar).Gametik damgalar nasıl kimliklendirilir? Bir damga, bir genin iki ebeveynsel kopyasını ayırt eden epigenetik modifikasyon olarak tanımlanabilir. Damga oluştuğunda, yazım düzeneğinin aynı çekirdek içinde annesel ve babasal gen kopyalarına farklı davranmasını da sağlamalıdır. Bir gametik damganın tüm gelişimsel evrelerde sürekli olarak varolması öngörülür (Fig. 3), dolayısıyla embriyonik veya yetişkin dokulardaki annesel ve babasal kromozomlar üzerindeki epigenetik modifikasyonlar karşılaştırılarak (Fig. 1'de özetlenen stratejiler kullanılarak) ve iki gametten birine kadar gelişimsel izleri sürülerek damgalar bulunabilir. Gametik damga adayları, DNA'yı kromozomlar şeklinde paketleyen histon proteinleri veya DNA modifikasyonları olabilir (Allis et al. 2014). Şu anda memelilerde bilinen iki çeşit epigenetik DNA modifikasyonu vardır: 5-metilsitozin ve 5-hidroksimetilsitozin (Li & Zhang 2014). Histonlar, metilasyon, asetilasyon, fosforilasyon, sumoilasyon ve ubikitilasyon da dahil olmak üzere çok çeşitli modifikasyonlar taşıyabilir (Allis et al. 2014). Ayrıca bunların yerini, spesifik işlevleri olan varyant histonlar alabilir (Henikoff & Smith 2014). Bu epigenetik modifikasyonların herhangi biri, bir damga olarak nitelendirilebilir. Bu epigenetik modifikasyonlardan sorumlu enzimlerin veya bir temel kofaktörün iki gametten birinde özel olarak ifade ediliyor olabileceği ve hücre bölündüğünde modifikasyonu kopyalamak için bir ebeveynsel kromozomla özel olarak ilişkili olduğu öngörülebilir. Ancak 3.bölümdeki "anahtar keşifler"de tanımlanacağı gibi, sadece 5-metilsitozinin memelilerdeki damgalanmış genler için gametik damga olarak işlediği açıkça gösterilmiştir ve şimdiye dek bilinen tek kalıtılabilen modifikasyondur.
Bir gametik damga, damgalanmış ifadeyi nasıl kontrol eder? Damganın nasıl işlediğini anlamak için üç parça bilgi gerekir: Damgayı hangi ebeveynsel kromozomun taşıdığı, damgalanmış genin ifade edilen allelini hangi ebeveynsel kromozomun taşıdığı ve damgalanmış genin ifade edilen veya susturulan alleline göre damgalanmış dizilimin konumu. Bu çeşit yaklaşımla, gametik damgaların bir seferde tüm bir gen kümesi üzerine etkiyebildiği ortaya konmuştur. Bu damgalanmış kümeler 3-12 damgalanmış gen içerir ve 100-3700 kb'lik genomik DNA kapsar (daha fazla ayrıntı için bkz. http://www.mousebook.org/catalog.php?catalog=imprinting). Herhangi bir kümedeki genlerin çoğu damgalanmış protein-kodlayan mRNA genleridir; ancak en az bir tanesi daima damgalanmış bir lncRNA'dır.
Bazı genlerin ebeveysel kromozomlardan birinden, bazılarının da diğerinden ifade edildiği, kümelerdeki damgalanmış genlerin düzenlenişi nedeniyle, damganın nasıl işlediğini belirlemek hiç de önemsiz değildir. Kümedeki tek bir genin üzerinde damganın etkisini incelemek olanaklıdır ama kümenin bütünü üzerinde damganın etkilerini incelemek daha bilgi verici olabilir. Bu daha ayrıntılı olarak 3.Bölüm'de tanımlanacaktır. Ancak net olan bir şey vardır. Doğa, gametlerden birinde damgalanmış bir geni önceden susturmak için damganın bir başlatıcıya doğru yöneltileceği en basit mekanizmayı seçmemiştir. Onun yerine, genel olarak, damgalar, çok sayıda genin ifadesini etkileyen ve aynı kromozom üzerinde uzak aralıklarla konumlanmış olan uzun-erimli cis-etkir düzenleyicilere doğru yöneltiliyor görünmektedir.
3. Genomik Damgalamada Anahtar Keşifler
3.1 Damgalanmış Genler Embriyonik ve Yenidoğansal Büyümeyi Kontrol Eder
Genomik damgalamanın memelilerdeki işlevi nedir? Bu soruyu yanıtlamanın bir yolu, bilinen damgalanmış genlerin in vivo (canlı içinde) işlevlerini belirlemektir. İşlev bozmak için "homolog rekombinasyon" tekniği kullanılarak gen diziliminin mutasyona uğratılmasıyla bu gerçekleştirilebilir. Bilinen damgalanmış genlerin çoğunun işlevi bu yolla belirlenmiştir (orijinal referanslar için bkz. http://www.mousebook.org/catalog.php?catalog=imprinting). Damgalanmış genler arasında en önemli işlevleri olanlar arasında embriyonun, plasentanın ve yenidoğanın büyümesini etkileyen genler sayılabilir. Bu kategoride, büyüme başlatıcılar olarak işleyen babasal ifadeli damgalanmış genler (Igf2, Peg1, Peg3, Rasgrf1, Dlk1) bulunur ve bu genleri kusurlu olan embriyolarda gelişim geriliği görülür. Ayrıca büyüme baskılayıcılar olarak işleyen annesel ifadeli damgalanmış genler de (Igf2r, Gnas, Cdkn1c, H19, Grb10) vardır ve bu genleri kusurlu olan embriyolarda büyüme artışı görülür. Bir diğer önemli kategoride ise davranışsal veya nörolojik kusurlu genler bulunur (örn. Nesp, Ube3a, Kcnq1). Bu sonuçlar bir açıdan umut kırıcıdır çünkü tüm damgalanmış genler için tek bir işlev tanımlamazlar. Yine de bu sonuçlar, damgalanmış genlerin büyük çoğunluğunun embriyonik veya yenidoğansal büyüme düzenleyiciler olarak işlediğini gösterir. Daha da ilginci, büyüme düzenleme becerisinin, annesel ifadeli büyüme düzenleyici genlerin yavrunun büyümesini baskılayıcı şekilde, bu kategorideki babasal ifadeli genlerin ise büyümeyi artırıcı şekilde etkiyecek biçimde düzgünce bölünmüş görünmesidir. Dahası, bazıları annesel davranışı değiştirerek yenidoğumsal büyüme hızını etkileyen sayısız sınanmış damgalanmış gen nörolojik süreçlerde etkindir.
3.2 Memelilerde Genomik Damgalamanın İşlevi
Gen işlevi çözümlemeleri, memelilerde genlerin neden damgalandığını anlamamıza yardım edebilir mi? Farklı memeli çeşitlerinde genomik damgalamanın değerlendirilmesi bilgi verici olmuştur. Fare ve insan gibi eteneli (plasentalı/ döleşli) memeliler ile opossum ve valabi gibi keseli memelilerde genomik damgalama bulunur. Platipus (ornitorenk) ve ekidne gibi yumurtlayan memeliler ise damgalanmış genlerden yoksun görünmektedir ancak geniş çaplı incelemeler henüz yapılmamıştır (Renfree et al. 2009). Eteneli ve keseli memeliler, embriyonun kendi büyümesi için kullanılacak annesel kaynakların miktarını doğrudan etkilemesini sağlayan bir üreme stratejisiyle yumurtlayan memelilerden ayrılır. Bunun tersine, yumurta içinde gelişen embriyoların annesel kaynakları doğrudan etkileme yetisi yoktur. Omurgasızların ve omurgalıların çoğu, yumurtlamalı bir üreme stratejisi kullanır. Ayrıca partenogenez de yapabildikleri dikkati çeker. Partenogenez, dişi gametin, erkek gametle döllenme olmaksızın, gelişerek yeni bir diploid birey olduğu bir üreme biçimidir (partenogenetik embriyolar aynı annesel genomun duplikasyonundan doğarken, Fig.2'de tanımlanan jinogenetik embriyoların iki farklı annesel genomdan doğduğuna dikkat edilmelidir). Organizmaların partenogenez yapabilme yetisinin, genomik damgalamanın hiç olmadığına işaret etmesi çok muhtemeldir çünkü babasal genomun zorunlu olmadığını gösterir. Memelilerde ise fetal büyümeyi damgalanmış gen ifadesinin kontrol etmesinin doğrudan bir sonucu, partenogenezin olanaklı olmamasıdır. Yaşayabilir yavrular üretmek için ebeveynlerin ikisine de gerek olması, memelilerin üremek için bütünüyle eşeyli üremeye bel bağlamasını zorunlu kılar (Fig. 4). Dolayısıyla aksi yönde iddialar olmasına karşın, partenogenez memelilerde şimdiye dek gözlemlenmemiştir ama Igf2 ve Dlk1 damgalanmış kümelerinin ifadesinin manipüle edilmesi, diploid annesel genomlu ender fareler üretmiştir (Kawahara et al. 2007).
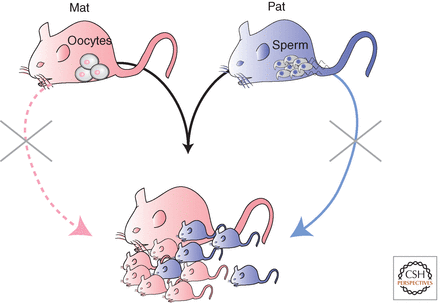 Figür-4. Damgalanmış genler memeli üremesinde rol oynar. Memeliler diploiddir ve üreme için haploid dişi yumurta ile haploid erkek spermin döllenmesi ve diploid embriyo oluşturması gerekir. Sadece dişiler anatomik olarak üreme donanımına sahiptir ama üremek için partenogenez kullanamazlar (pembe kesikli çizgi ile temsil edilen olasılık) çünkü fetal gelişim için gereken damgalanmış genler, annesel kromozomlarda damgalanmış ve susturulmuştur. Bu genler sadece babasal kromozomlardan ifade edilebilir; dolayısıyla memelilerde üreme için her iki ebeveynsel kromozom gereklidir. Partenogenez, aynı annesel genomun iki kopyasından diploid yavru üretilmesidir.
Figür-4. Damgalanmış genler memeli üremesinde rol oynar. Memeliler diploiddir ve üreme için haploid dişi yumurta ile haploid erkek spermin döllenmesi ve diploid embriyo oluşturması gerekir. Sadece dişiler anatomik olarak üreme donanımına sahiptir ama üremek için partenogenez kullanamazlar (pembe kesikli çizgi ile temsil edilen olasılık) çünkü fetal gelişim için gereken damgalanmış genler, annesel kromozomlarda damgalanmış ve susturulmuştur. Bu genler sadece babasal kromozomlardan ifade edilebilir; dolayısıyla memelilerde üreme için her iki ebeveynsel kromozom gereklidir. Partenogenez, aynı annesel genomun iki kopyasından diploid yavru üretilmesidir.Neden genomik damgalamanın genel olarak tüm omurgalılarda değil de, bazı memelilerde evrilmesi gerekmiştir? Genomik damgalamanın üç özelliği, eşdeğer çekicilikte iki hipoteze uyan kanıtlar sağlar: Çoğu damgalanmış genin büyüme düzenleyici işlevi, damgalanmış genlerin eteneli ve keseli memelilerle sınırlanması, fetal gelişim için babasal genomun gerekliliği.
İlk hipotez, genomik damgalamanın "ebeveynsel çekişme" durumuna yanıt olarak evrildiğini ileri sürer (Moore & Haig 1991). Bu, annesel genom ile babasal genomun çıkarlarının ters düşmesinden doğar: Embriyonik büyüme tek bir ebeveyne bağlıdır, ama genomu iki ebeveynden gelen bir embriyo tarafından etki altına alınır. Babasal ifadeli damgalanmış genler embriyonik büyümeyi artırmaya ve böylece belli bir babasal genomu taşıyan yavrunun gürbüzlüğünü yükseltmeye çalışır. Annesel ifadeli damgalanmış genler ise fetal büyümeyi baskılamaya çalışır. Bu, annesel kaynakların tüm yavrulara daha eşitçe dağılmasını ve farklı babasal genomlar taşıyor olabilecek çok sayıda yavruya annesel genomun aktarımını artırır.
İkinci hipotez "trofoblast savunma" olarak adlandırılır (Varmuza & Mann 1994). Bu da, annesel genomun, içsel üreme için anatomik olarak donanımlı olmanın olası spantane oosit aktivasyonunun tam embriyonik gelişime yol açması gibi sonuçlarından dolayı risk altında olduğunu ileri sürer. Erkekler içsel üreme için gereken anatomik donanımdan yoksun olduğu için spermatozoanın spontane aktivasyonunun getirebileceği bir risk yoktur. Dolayısıyla damgalama, ya annesel genomdaki plasental gelişim başlatıcı genleri susturmalı ya da süreci sınırlayan genleri etkinleştirmelidir. Annesel rahim damarlarının plasenta tarafından işgali için gereken genler, sonuç olarak sadece döllenmeden sonra bir babasal genomdan ifade edilir.
Bu hipotezlerden birisi memelilerde genomik damgalamanın evrimini açıklar mı; evetse hangisi? Hipotezlerin her ikisi de damgalanmış genlerin, plasenta gelişim ve işlevinin düzenlenmesinde rolü olduğuna işaret eder ancak ne ebeveynsel çekişme ne de trofoblast savunma modeli, verilerin tümü için tam bir açıklama vermez (Wilkins & Haig 2003). Damgalanmış genlerin bitki endosperminde de tanımlanmış olması ilginçtir; endosperm, plasenta ile karşılaştırılabilecek bir dokudur çünkü ebeveyn bitkiden embriyoya besin kaynaklarının aktarımını yapar (Grossniklaus & Paro 2014). Bu bulgu, genomik damgalamanın ebeveyn ile yavru arasında besin aktarımını düzenlemek için bir yol olarak evrildiği iddiasını güçlendirir ama nedenini anlatmaz.
Memelilerde genomik damgalamanın daha eksiksiz veya alternatif açıklamaları, iki kaynaktan gelebilir. İlki, tek bir damgalanmış gen üründen yoksun farelerin fenotipini araştırmanın tersine, tüm bir gen kümesi boyunca "damgalama"nın işlevini araştırmak olabilir. Bunun için bir damgayı geri alma ve tüm damgalanmış küme boyunca iki-ebeveynsel (İng. biparental) gen ifadesi üretme yetisi gerekir. İkinci yaklaşım, tam olarak hangi genlerin damgalanmış olduğunu öğrenmektir. Bir kümedeki genlerin hepsinin damgalama mekanizmasının kasıtlı hedefleri olmayıp, bazılarının sadece sürecin "masum seyirciler"i olması, işlevlerinin genomik damgalamanın rolü hakkında bilgi verici olmaması olasıdır. Damgalama mekanizması tarafından etkilenen masum seyirci genlerin varlığı, gelişimde net bir biyolojik işlevi olmayan damgalanmış genlerin merak uyandırıcı bolluğunu tatminkâr biçimde açıklayabilir.
3.3 Damgalanmış Genler, Damga Kontrol Elemanları Tarafından Kontrol Edilir ve Kümelenmiştir
Şimdiye dek, X kromozomu da dahil olmak üzere 17 fare kromozomuna 150 civarında damgalanmış gen haritalandı. Tanımlanan damgalanmış genlerin %80'den fazlası, iki ya da daha fazla gen içeren 16 genomik bölgeye kümelenmişti (Wan & Bartolomei 2008). Damgalanmış genlerin kümelerinin keşfi, ortak bir DNA elemanının cis'teki çok sayıda genin damgalanmış ifadesini düzenliyor olabileceğinin güçlü bir işaretiydi. Şimdiye dek, 16 damgalanmış kümeden yedi tanesi iyi bir şekilde karakterize edildi (özellikleri belirlendi); bunlar kümedeki birincil damgalanmış mRNA geninin veya ilişkilendirilen hastalığın adına göre Tablo.1'de listeleniyor (örn. Prader-Willi sendromu için Pws kümesi; Beaudet & Zoghbi 2014). Bu yedi küme üç ilâ 12 (veya daha fazla) damgalanmış gen içeriyor ve 80-3700 kb'lık DNA'ya yayılıyor.
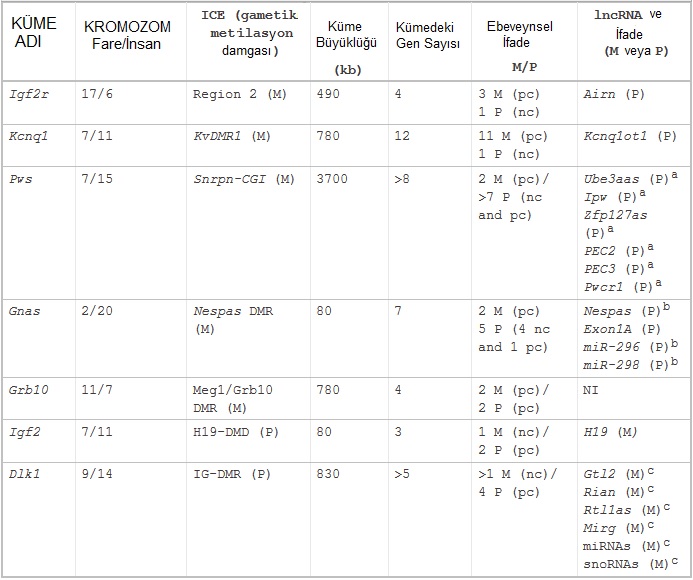 Tablo-1. Fare genomundaki damgalanmış gen kümelerinin özellikleri. Küme büyüklüğünün ve kümedeki gen sayısının netleşmemiş olduğunu ve damgalanmış ifadenin genom çapında çözümlemesini beklediğine dikkat ediniz. Pws ve Dlk1 kümeleri, ayrık genlerin sayısı henüz netleşmemiş olan çakışan yazımlar içerir. Ayrıntılar metinde verilmiştir. M, annesel (İng. maternal); P, babasal (İng. paternal); DMR, farklı metillenmiş bölge (İng. differentially methylated region); pc, protein kodlayan (İng. protein coding); nc, kodlamayan (İng. noncoding) RNA; NI, tanımlanmamış (İng. none identified); miRNA, mikro (İng. micro) RNA; snoRNA, küçük çekirdekçiksel (İng. small nucleolar) RNA.
Tablo-1. Fare genomundaki damgalanmış gen kümelerinin özellikleri. Küme büyüklüğünün ve kümedeki gen sayısının netleşmemiş olduğunu ve damgalanmış ifadenin genom çapında çözümlemesini beklediğine dikkat ediniz. Pws ve Dlk1 kümeleri, ayrık genlerin sayısı henüz netleşmemiş olan çakışan yazımlar içerir. Ayrıntılar metinde verilmiştir. M, annesel (İng. maternal); P, babasal (İng. paternal); DMR, farklı metillenmiş bölge (İng. differentially methylated region); pc, protein kodlayan (İng. protein coding); nc, kodlamayan (İng. noncoding) RNA; NI, tanımlanmamış (İng. none identified); miRNA, mikro (İng. micro) RNA; snoRNA, küçük çekirdekçiksel (İng. small nucleolar) RNA.a Tek bir uzun lncRNA olabilir.
b Nespas yazımının parçası.
c Tek veya çok sayıda lncRNA'nın parçası olabilir. Tablo-1. Fare genomundaki damgalanmış gen kümelerinin özellikleri. Küme büyüklüğünün ve kümedeki gen sayısının netleşmemiş olduğunu ve damgalanmış ifadenin genom çapında çözümlemesini beklediğine dikkat ediniz. Pws ve Dlk1 kümeleri, ayrık genlerin sayısı henüz netleşmemiş olan çakışan yazımlar içerir. Ayrıntılar metinde verilmiştir. M, annesel (İng. maternal); P, babasal (İng. paternal); DMR, farklı metillenmiş bölge (İng. differentially methylated region); pc, protein kodlayan (İng. protein coding); nc, kodlamayan (İng. noncoding) RNA; NI, tanımlanmamış (İng. none identified); miRNA, mikro (İng. micro) RNA; snoRNA, küçük çekirdekçiksel (İng. small nucleolar) RNA.a Tek bir uzun lncRNA olabilir.b Nespas yazımının parçası.c Tek veya çok sayıda lncRNA'nın parçası olabilir.
Bu yedi kümenin bir ortak özelliği, gametik DMR (farklı şekilde DNA-metillenmiş bölge; İng. differentially DNA-methylated region) olarak bilinen bir gametik metilasyon damgası taşıyan bir DNA diziliminin varlığıdır. Bir gametik DNA metilasyon damgası, gametlerden birinde bulunan ve embriyonun diploid hücrelerindeki sadece bir ebeveynsel kromozom üzerinde sürdürülen bir metilasyon damgası olarak tanımlanır. Gametik DMR, beş kümede (Igf2r, Kcnq1, Gnas, Grb10 ve Pws) oojenezde edinilmiş bir annesel metilasyon damgası taşırken, iki kümede (Igf2 ve Dlk1) spermatojenez sırasında edinilmiş babasal bir metilasyon damgası taşır. Bu örneklerde, gametik DMR kümenin bütününün veya bir kısmının damgalanmış ifadesini kontrol eder ve dolayısıyla kümenin damga kontrol elemanı (İng. imprint control element - ICE) olarak belirtilir (Barlow 2011).
Tablo.1'de her bir damgalanmış gen kümesinin çok sayıda mRNA ve Grb10 istisnasıyla birlikte en az bir lncRNA içerdiği görülür. İki trend belirir. Birincisi, her bir kümedeki damgalanmış protein-kodlayan genler büyük ölçüde aynı ebeveynsel kromozomdan ifade edilirken, lncRNA'lar karşıt ebeveynsel kromozomdan ifade edilir (Fig.5'te bir annesel gametik DMR için gösterildiği gibi). İkincisi, ICE delesyonu sadece lncRNA'yı ifade eden ebeveynsel allelden silindiği zaman damgalanmış ifade yitimine neden olur.
Tablo.1'de üç kümede (Igf2r, Kcnq1 ve Gnas) lncRNA başlatıcı damgalanmış mRNA'lardan birinin bir intronunda bulunurken, geri kalan kümelerde lncRNA başlatıcının ayrı olduğu ama damgalanmış mRNA genlerine yakın bulunduğu görülür. Bir damgalanmış kümedeki etkin ve suskun genlerin bu yakın iç-içeliği, damgalanmış genleri etkileyen susturma ve etkinleştirme mekanizmalarının etrafa yayılmadığına ve etkiledikleri genle sınırlı olabileceklerine işaret eder. Özellikle belirtmek gerekirse, bir suskun lncRNA'nın başlatıcısının, etkin olarak yazımı yapılan bir genin intronunda bulunabilmesi, susturma mekanizmalarının bir genin uzunluğu oyunca bile yayılmış olmayavileceğine işaret eder; sadece düzenleyici elemanlarla sınırlı olabilir.
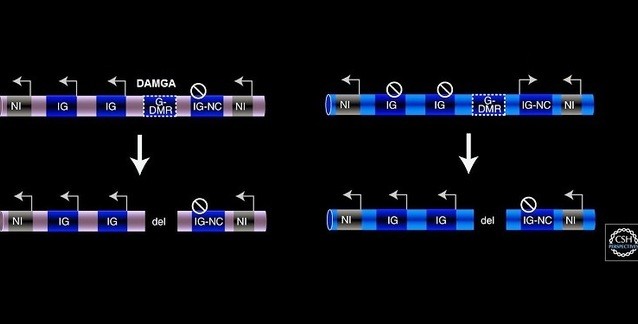
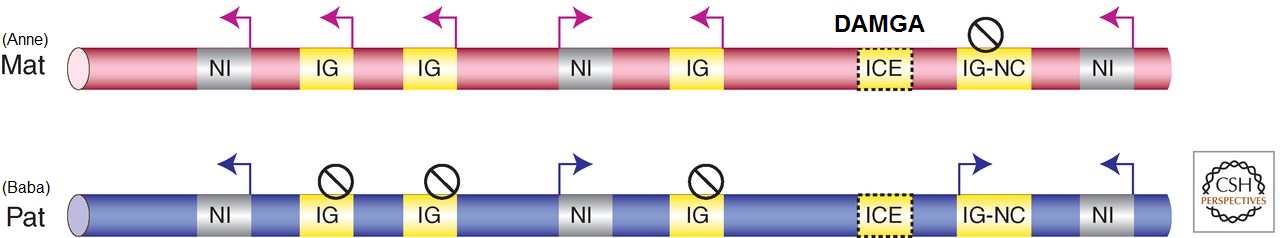 Figür-5. Damgalanmış genler, ebeveynsel allellerin birinden ifade edilir ve çoğunlukla kümelenmişlerdir. Çoğu damgalanmış gen (sarı), çok sayıda protein kodlayan mRNA (IG) ve en az bir kodlamayan RNA (IG-NC) içeren kümeler hâlinde bulunur. Damgalanmamış genler de mevcut olabilir (gri NI). Damgalama mekanizması cis etkirdir ve damgalanmış ifade ebeveynsel gametlerden birinden kalıtılmış bir epigenetik damga taşıyan bir damga komtrol elemanı (ICE) tarafından kontrol edilir. Bir çift fiploid kromozom gösterilmiştir: Pembe olan annesel kökenli, mavi olan babasal kökenlidir. Ok, ifade edilen gen; çizgili daire, baskılanmış gen.
Figür-5. Damgalanmış genler, ebeveynsel allellerin birinden ifade edilir ve çoğunlukla kümelenmişlerdir. Çoğu damgalanmış gen (sarı), çok sayıda protein kodlayan mRNA (IG) ve en az bir kodlamayan RNA (IG-NC) içeren kümeler hâlinde bulunur. Damgalanmamış genler de mevcut olabilir (gri NI). Damgalama mekanizması cis etkirdir ve damgalanmış ifade ebeveynsel gametlerden birinden kalıtılmış bir epigenetik damga taşıyan bir damga komtrol elemanı (ICE) tarafından kontrol edilir. Bir çift fiploid kromozom gösterilmiştir: Pembe olan annesel kökenli, mavi olan babasal kökenlidir. Ok, ifade edilen gen; çizgili daire, baskılanmış gen.Gametik DMR'nin rolü nedir? Gametik DMR'lerin annesel veya babasal metillenmiş olabilmeleri gerçeğine rağmen, bu elemanları silen deneyler geniş ölçüde benzer sonuçlar üretmiştir ama birkaç ilginç istisna da vardır (Fig. 6). Üç kümede (Igf2r, Kcnq1, Dlk1) metillenmiş gametik DMR'nin deneysel silinmesi hiçbir etki üretmez. Tersine, metillenmemiş gametik DMR'nin silinmesi, ebeveyne-özgü ifadeyi ortadan kaldırarak, cis'te lncRNA ifadesinin ve biallellik mRNA ifadesinin yitimine neden olur (Lin et al. 1995; Zwart et al. 2001; Fitzpatrick et al. 2002). İki küme (Gnas ve Pws) birden fazla gametik DMR içeriyor gibi görünür ve daha karmaşık bir davranış gösterir; yine de Fig.6'da sunulan örüntü ile bazı benzerlikler paylaşır (Williamson et al. 2006). Ancak Igf2 kümesi farklı davranır: Hem metillenmiş hem de metillenmemiş gametik DMR'nin ikisinin de silinmesi, cis'teki mRNA ve lncRNA ifadesinde değişimlere neden olur (Thorvaldsen et al. 1998).
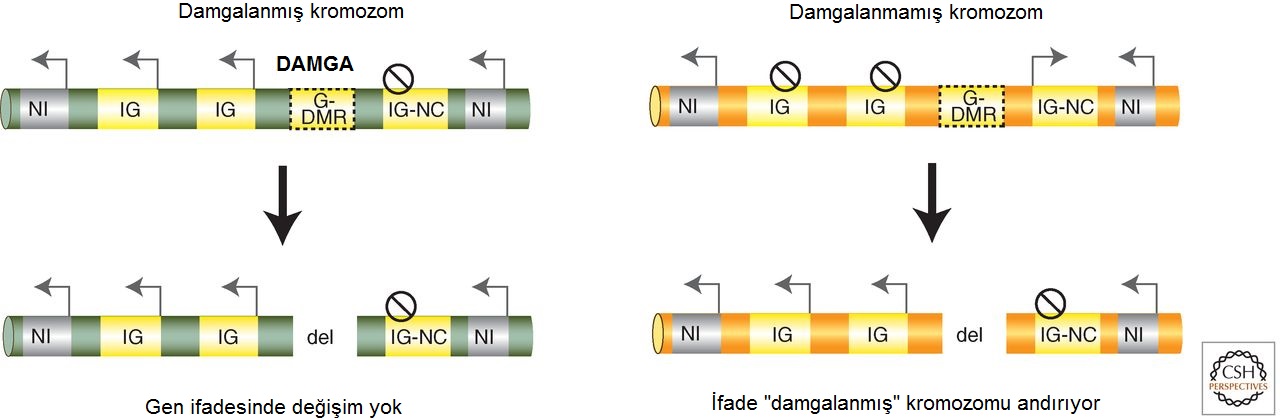 Figür-6. Damgalanmış ifade gametik DMR'ler (G-DMR) tarafından düzenlenir. (Solda) Damgalanmış kromozomdan gametik DMR'nin silinmesinin etkisi. (Sağda) Damgalanmamış kromozomdan G-DMR'nin silinmesinin etkisi (sarı). Çoğu damgalanmış kümede (örn. Igf2r, Kcnq1 ve Dlk1), G-DMR'nin deneysel silinmesi sadece damgalanmamış G-DMR taşıyan kromozomu etkiler. Bu durum, damgalanmış protein-kodlayan mRNA genlerinin (IG) baskılanmasının kaybıyla ve damgalanmış lncRNA geninin (IG-NC) baskılanmasının kazanımıyla sonuçlanır. Burada gösterilmeyen bazı damgalanmış kümelerde (Igf2 ve Pws), metillenmiş G-DMR'nin de cis'teki damgalanmış mRNA'ların bazılarının ifadesi için gerekli göründüğüne dikkat ediniz. (del, silinmiş (İng. deleted) DNA; G-DMR, gametik farklı DNA-metillenmiş bölge; NG, damgalanmamış gen; ok, ifade edilen allel; çizgili daire, baskılanmış allel; damga, cis'teki gen ifadesinde bir değişime yol açan epigenetik modifikasyon.
Figür-6. Damgalanmış ifade gametik DMR'ler (G-DMR) tarafından düzenlenir. (Solda) Damgalanmış kromozomdan gametik DMR'nin silinmesinin etkisi. (Sağda) Damgalanmamış kromozomdan G-DMR'nin silinmesinin etkisi (sarı). Çoğu damgalanmış kümede (örn. Igf2r, Kcnq1 ve Dlk1), G-DMR'nin deneysel silinmesi sadece damgalanmamış G-DMR taşıyan kromozomu etkiler. Bu durum, damgalanmış protein-kodlayan mRNA genlerinin (IG) baskılanmasının kaybıyla ve damgalanmış lncRNA geninin (IG-NC) baskılanmasının kazanımıyla sonuçlanır. Burada gösterilmeyen bazı damgalanmış kümelerde (Igf2 ve Pws), metillenmiş G-DMR'nin de cis'teki damgalanmış mRNA'ların bazılarının ifadesi için gerekli göründüğüne dikkat ediniz. (del, silinmiş (İng. deleted) DNA; G-DMR, gametik farklı DNA-metillenmiş bölge; NG, damgalanmamış gen; ok, ifade edilen allel; çizgili daire, baskılanmış allel; damga, cis'teki gen ifadesinde bir değişime yol açan epigenetik modifikasyon.Yukarıdaki gametik DMR silme deneylerinden çıkan sonuçlar, ilk bakışta gametik DMR'ler için ortak bir işleve işaret etmez. Ancak tam işlevlerini anlamak, her bir kümedeki damgalanmış genlere göre DMR'nin konumunu bilmeye bağlıdır. En basit örüntülü üç kümede (Igf2r, Kcnq1 ve Dlk1), gametik DMR, lncRNA'nın ifadesini ya içerir ya da kontrol eder; dolayısıyla bu elemanın silinmesinin lncRNA ifadesinin yitimine yol açacağı nettir. Igf2 kümesindeki gametik DMR ise H19 transkripsiyonunu (yazımını) doğrudan başlatmaz ama Igf2 ve H19 ve onların ortak yükselticileri arasındaki etkileşimi değiştirir ve bu yolla ifadelerini düzenler. Bu farklara rağmen, genel olarak, metillenmemiş gametik DMR'nin altı kümenin hepsinde lncRNA ifadesinin bir pozitif düzenleyicisi olduğu sezilir ve DNA metillenmesi damgasının varlığı, lncRNA'nın baskılanması ile ilişkilendirilir. Gametik DMR'lerin silinmesinden elde edilen verilerden çıkan sonuç, bu bölgeleri, etkinlikleri DNA metilasyonu tarafından düzenlenen ICE olarak açıkça tanımlar.
3.4 Damgalanmış Gen Kümeleri En Az Bir Tane lncRNA İçerir
Damgalanmış kümelerin çoğunluğu, şu anda >200 nükleotidli kodlamayan bir yazım (transkript) olarak tanımlanan bir lncRNA içerir (Guttman & Rinn 2012). Eskiden lncRNA'ların, RNA işleme ve çevrimi (translasyon) ile ilgili olanlar dışında, memeli genomunda ender rastlanan bir şey olduğu düşünülüyordu. Artık fare ve insan genom dizilimlerinin erişilebilir olması nedeniyle, transkriptom analizleri, verilen bir hücre popülasyonundaki tüm RNA yazımlarının (transkriptlerinin) tanımlanmasıyla sonuçlandı. Bu çalışma, memeli transkriptomunun büyük bir bölümünün, lncRNA'lardan oluştuğunu gösterdi. Gen düzenleme işlevleri taşıyan çeşitli tiplerde memeli kodlamayan RNA'ları (ncRNA) vardır; bunlar arasında susturma yolaklarına katılan "kısa" ncRNA'lar (Martienssen & Moazed 2014), X kromozomu aktivasyonunda veya inaktivasyonunda görev alan Xist gibi "daha uzun" işlenmiş lncRNA'lar (Brockdorff & Turner 2014: Memelilerde Dozaj Telafisi; Kuroda & Lucchesi 2014) ve protein-kodlayan genlerin cis veya trans aktivasyonu veya susturulmasıyla ilişkilendirilen lncRNA'lar (Allis et al. 2014; Rinn 2014) bulunur.
Damgalanmış gen kümeleriyle hangi çeşit lncRNA'lar ilişkilendirilir? Tablo-1'de gösterilen iyi bir şekilde karakterize edilmiş (özellikleri belirlenmiş) damgalanmış genler ile ilişkilendirilen lncRNA'larn çözümlemesi hâlâ eksik olup, bazı benzerleriklerle birlikte bazı farklar da göze çarpar. Üç damgalanmış lncRNA, alışılmadık ölçüde uzun olgun RNA'lardır: Airn 108 kb'dır (Lyle et al. 2000), Kcnq1ot1 ∼100 kb'dır (Pauler et al. 2012) ve Ube3aas >1000 kb olabilir (Landers et al. 2004). Tersine, H19 lncRNA sadece 2.3 kb'dır (Brannan et al. 1990). Gtl2 lncRNA çok sayıda alternatif biçimde uçbirleştirmiş (splays yapmış; İng. spliced) yazımlar (transkriptler) içerir; ancak, aşağı yönlü (İng. downstream) interjenik yazım da (transkripsiyon) fark edilmiş ve daha uzun yazım birimlerinin de olası olduğuna işaret etmiştir (Tierling et al. 2005). Nespas lncRNA, RNA lekelerinin üzerinde çözülemeyecek kadar iridir ve tam boyutu 27 kb'ı aşar (Robson et al. 2012). Bu sonuncu lncRNA'lar, düşük bir intron-ekson oranıyla intron fakiri gibi görünür veya olgun yazımlar (transkriptler) olarak uçbirleştirmemiştir (İng. unspliced) (Seidl et al. 2006; Pandey et al. 2008). Bir diğer özellik de, üç damgalanmış lncRNA'nın (H19, Ube3aas ve Gtl2 aşağı yönlü yazımlar) snoRNA'lar (İng. small nucleolar RNA; küçük çekirdekçiksel RNA'lar ribozomal DNA, snRNA ve muhtemelen mRNA modifikasyonlarını yöneterek, yazım-sonrası (post-transkripsiyon) düzenleyiciler olarak görev yapar) ve miRNA'lar (mikro RNA) için konak yazımlar olarak davranmasıdır. Küçük çekirdekçiksel RNA'lar (snoRNA), kümedeki damgalanmış mRNA genlerine doğru yönelmemiştir ve damgalama mekanizmasının kendisinde bir rol oynayıp oynamadıkları şu anda net değildir (Seitz et al. 2004). Benzer biçimde, Gtl2 lncRNA'lar ve H19'daki miRNA'lar, mRNA genlerinin yazım-sonrası baskılanmasıyla ilgilidir ama kümenin damgalanmış ifadesini düzenlemede doğrudan bir rol oynamaz (Davis et al. 2005; Keniry et al. 2012). Damgalanmış lncRNA'ların iki özelliği, onların kümedeki damgalanmış mRNA (yani protein-kodlayan) genlerin susturulmasında rol oynayabileceğine işaret eder. Birincisi, lncRNA genellikle damgalanmış mRNA genlerine kıyasla karşıt ebeveyne-özgü ifade sergiler (Tablo 1). İkincisi, kümenin bütününün damgalanmış ifadesini kontrol eden gametik metilasyon damgası taşıyan DMR, çok kez lncRNA başlatıcı ile çakışır (Airn region2, KvDMR1, Snrpn-CGI ve Nespas-DMR). Bu bulgu, damgaların, her bir damgalanmış kümedeki lncRNA'yı düzenlemek için evrildiğine işaret ediyor olabilir. Bu yorum, Igf2r, Kcnq1, Gnas, Pws ve Dlk1 kümelerinde test edildiği üzere, gametik DMR taşıyan metillenmemiş dizilimi silen deneylerin lncRNA ifadesinin yitimi ile beraber gelen damgalanmış mRNA genlerinin (Fig. 6) ifadesindeki kazanca neden olması tarafından desteklenir (Wutz et al. 1997; Bielinska et al. 2000; Fitzpatrick et al. 2002; Lin et al. 2003; Williamson et al. 2006).
Şu anda lncRNA'nın kendisinin rolünü doğrudan sınayan deneyler, çok sayıda lokus (Airn, Nespas, Kcnq1ot1 ve H19 lncRNA'lar) için gerçekleştirilmiş durumdadır. Bu lncRNA'lar, içsel (endojen) lokusun genetik manipülasyonu ile çözümlenmiştir. İlk üç lokus, lncRNA'yı kırpmak için bir poliadenilasyon sinyali yerleştirilerek denenmiştir. 108 kb'lık Airn lncRNA'nın 3 kb'a kırpılması, Igf2r kümesindeki üç mRNA geninin hepsini susturmak için lncRNA'nın kendisinin gerekli olduğunu göstererek, bu lncRNA için açık bir düzenleyici role işaret etmiştir (Sleutels et al. 2002). Yaklaşık 100 kb'lık Kcnq1ot1 lncRNA'nın 1.5 kb'a kırpılması da, daha büyük Kcnq1 kümesindeki 10 mRNA geninin hepsinin susturulması için bu lncRNA'nın doğrudan gerektiğini göstermiştir (Mancini-DiNardo et al. 2006). Son olarak, yaklaşık 27 kb'lık Nespas lncRNA'nın kırpılması, Gnas damgalanmış kümesindeki çakışan Nesp genini susturmak için onun gerekli olduğunu göstermiştir (Williamson et al. 2011). Tersine, H19 ncRNA'nın tümden silinmesinin, endoderm dokulardaki Igf2 kümesinde bulunan damga üzerinde bir etkisi yoktur ama mezoderm dokuda bazı damga yitimleri görülmüştür (Schmidt et al. 1999). Dolayısıyla, üç annesel damgalı küme (Igf2r, Kcnq1 ve Gnas), ortak bir lncRNA'ya bağlı susturma mekanizması paylaşırken, şimdiye dek incelenen babasal damgalı tek küme (Igf2), farklı bir yalıtıcıya bağlı model kullanır (bkz. Bölüm 3.6).
3.5 Genomik Damgalamada DNA Metilasyonunun Rolü
İlk üç içsel (endojen) damgalanmış genin 1991 yılında tanımlanması, hücrenin epigenetik mekanizmasının damgalanmış bir geni ebeveynsel kimliğiyle nasıl işaretlediğini, araştırmacıların incelemesine olanak tanıdı. İlk ve en kolay sınanabilecek aday DNA metilasyonu idi; DNA metilasyonu, memelilerde CpG dinükleotidlerindeki sitozin rezidüsüne kovalent olarak bir metil grubu ekleyen bir modifikasyondur. DNA metilasyonu, de novo (yeni) metiltransferazların eylemi yoluyla edinilir ve koruma metiltransferazlarının eylemiyle hücre her bölündüğünde in situ (hücre içinde) korunur (bkz. Li & Zhang 2014). Dolayısıyla bu modifikasyon, Fig.3'te belirtilen, bir ebeveynsel kimlik işareti yani "damga"da olması gereken ölçütleri sağlar çünkü (1) sadece tek bir gamette etkiyen de novo metiltransferazlar tarafından ya spermde ya da oositte ayarlanır, (2) bir korunum metiltransferazı tarafından her bir embriyonik hücre bölünmesinde sabit şekilde yayılabilir, (3) ya pasif demetilasyonla (metilasyon korunumunu sağlamada başarısızlığın izlediği DNA eşlenmesi) ya da demetilleyici etkinliğin eylemi yoluyla (muhtemelen on-onbir enzim translokasyon ailesi tarafından 5-metilsitozinin 5-hidroksimetilsitozine dönüşümü yoluyla veya DNA onarım mekanizması tarafından 5-metilsitozinin atılması yoluyla ) bir sonraki kuşakta damgayı yeni baştan ayarlamak için germ hattında silinebilir.
DNA metilasyonunun, genomik damgalamada iki farklı işlev gerçekleştirme potansiyeli vardır. Sadece gametlerden birindeki kromozomlar tarafından de novo edinilerek, damgalama işareti görevi görebilir. Ayrıca ebeveynsel allellerden birini susturabilir çünkü DNA metilasyonu gen baskılanması ile ilişkilendirilir (Li & Zhang 2014). Hangi işleve sahip olduğunu belirlemek için önce DNA metilasyonunun sadece bir ebeveynsel kromozomda olduğunu (yani bir DMR olduğunu) göstermek gerekir. İkinci olarak, kümedeki hangi damgalanmış genin ve hangi düzenleyici dizilimlerin DNA metilasyonu tarafından işaretlendiğini tanımlamak gerekir. Bir başlatıcı üzerindeki veya uzak pozitif ya da negatif düzenleyici elemanlar üzerindeki metilasyon işaretlerinin konumu, gen ifadesi için farklı sonuçlar doğurur. Son olarak, gelişim sırasında DMR'nin ne zaman oluştuğunu belirlemek gerekir. Eğer gametojenez sırasında oluşuyor ve somatik hücrelerde sürekli olarak yeri korunuyorsa (gametik DMR olarak bilinir), damgalama işareti görevi görüyor olabilir. Ancak eğer ebeveynsel kromozomların ikisi de aynı hücre içindeyken yani embriyo diploid hâle geldikten sonra gen üzerine yerleştiriliyorsa (somatik DMR olarak bilinir), kimlik işareti görevi görmesi olasılık dışıdır ama ebeveyne-özgü susturmayı sürdürme görevi görebilir.
Ebeveynsel allele-özgü DNA metilasyonu, incelenen damgalanmış kümelerin çoğunda bulunmuştur. Örneğin, Igf2 kümesi, sadece babasal gamette metillenmiş olan ve sonrasında tüm somatik dokularda sürdürülen, H19 lncRNA başlatıcıdan 2 kb yukarı yönde (İng. upstream) konumlanan bir gametik DMR'ye sahiptir (Bartolomei et al. 1993). Benzer bir gametik DMR, Airn lncRNA'nın başlatıcısını kaplıyor olarak, sadece sessiz annesel gen kopyasında varolduğu ve dişi gamette edinildiği tanımlanmıştır (Stoger et al. 1993). Şaşırtıcı bir şekilde, bu kümelerdeki (sırasıyla, Igf2 ve Igf2r) birincil damgalanmış protein-kodlayan genlerin başlatıcılarında gametik DMR'ler belirlenmemiştir. Sessiz Igf2 başlatıcısında DNA metilasyonu yokken, sessiz Igf2r başlatıcısı döllenmeden sonra yerleştirilen bir somatik DMR içinde yer alır (Sasaki et al. 1992; Stoger et al. 1993). Damgalanmış lncRNA'nın sessiz kopyasını taşıyan kromozom üzerindeki gametik DMR'lere (bkz. Fig.6) ilişkin benzer bulgulara, Pws, Kcnq1, Gnas, Dlk1 ve Grb10 dahil olmak üzere diğer iyi incelenmiş gen kümelerinde de rastlanmıştır (Shemer et al. 1997; Liu et al. 2000; Takada et al. 2002; Yatsuki et al. 2002; Shiura et al. 2009).
Somatik DMR'ler nispeten enderdir ama bazı damgalanmış kümeler için raporlanmıştır ki bu da bu epigenetik modifikasyon çeşidinin damgalanmış gen ifadesinin sürdürülmesinde sınırlı bir rolünün olduğuna işaret eder (Stoger et al. 1993; Moore et al. 1997; Yatsuki et al. 2002; John & Lefebvre, 2011). Farelerde gametik DMR'lerin silinmesi, çok sayıda gen için damgalamanın tümüyle yitirilmesi sonucunu doğurarak, bu DMR'ler sınıfının, kümenin bütünü için başlıca ICE'lerden biri olarak da görev yaptığını kanıtlar (Fig.6) (Wutz et al. 1997; Thorvaldsen et al. 1998; Bielinska et al. 2000; Fitzpatrick et al. 2002; Lin et al. 2003; Williamson et al. 2006). Tersine, somatik DMR'lerin silinmesi, komşusu olan damgalanmış genin ifadesini etkiler ama kümedeki diğer genlerde damgalanmış ifade sürdürülür (Constancia et al. 2000; Sleutels et al. 2003).
Dnmt gen ailesindeki mutasyonların neden olduğu, DNA metilasyonun genom çapında kusurlu olma durumu, damgalnmış gen ifadesinin düzenlenmesinde DNA metilasyonunun temel rolünün altını çizer. Mutasyona uğrayan de novo DNA metiltransferaz Dnmt3a, DNA metiltransferaz uyarım faktörü Dmnt3L veya Dnmt1 sürdürücü DNA metiltransferaz, damgalanmış gen ifadesinde değişiklikler sergileyen DNA metilasyonu kusurlu embriyolar üretir. Dört damgalanmış küme (Igf2, Igf2r, Kcnq1 ve Dlk1) için gösterilen pertürbasyonların çeşidi, DNA metilasyonunun genellikle gametik DMR'nin eylemini baskılayıcı davrandığına işaret eder. Dolayısıyla, DNA metilasyonunun yokluğunda, gametik DMR düzgün işleyemez (yani lncRNA'yı susturamaz). Bunun sonucunda, lncRNA düzensiz bir şekilde ifade edilir ve Igf2, Igf2r, Kcnq1 ve Dlk1 dahil olmak üzere çeşitli damgalanmış protein-kodlayan genler, her iki ebeveynsel kromozomda da baskılanmış duruma gelir. Bu da, bu mRNA genlerinin ifade edilmesi için bir cis-düzenleyici elemanın epigenetik modifikasyonunun gerektiğine işaret eder. Dikkat çekici bir şekilde, normalde sadece metillenmemiş gametik DMR'yi taşıyan kromozom üzerinde ifade edilen H19 lncRNA, her iki ebeveynsel kromozomda da ifade edillir duruma gelir. Bu genel örüntünün bazı istisnaları, sadece plasentada damgalanmış ifade sergileyen genler için raporlanmıştır (Lewis et al. 2004).
Gametik damga olarak kullanılan başka epigenetik modifikasyon çeşitleri var mıdır? Memeli genomundaki genetik bilgiyi modifiye edecek şekilde davranan epigenetik mekanizmaların sırf bolluğuna bakılırsa, DNA metilasyonunun yegâne damgalama mekanizması olması pek muhtemel değildir. Kromatin etkinlik durumlarını etkileyen histon modifikasyonları da olası ebeveynsel damga adaylarıdır çünkü Fig.3'te belirtilen önkoşulların çoğunu sağlarlar. Örneklerden birinde, EED olarak bilinen Polycomb grubu proteinin (H3K27'nin yani lizin 27'deki H3 histonunun metilasyonunu katalizleyen PRC2 kompleksinin parçası) plasentadaki birkaç babasal baskılanmış geni etkilediği gösterilmiştir. Eed mutasyonun genomik damgalama üzerindeki etkileri ise DNA metilasyonununkine kıyasla nispeten küçüktür (Mager et al. 2003). Bir diğer örnekte, yine sadece plasentada, birkaç damgalanmış geni baskılamak için spesifik olarak H3K9 üzerine etkiyen EHMT2 histon metiltransferaz gerekir (Nagano et al. 2008). Dolayısıyla, şu âna kadarki veriler, histon modifikasyonlarının ve modifiye edici enzimlerin, genomik damgalamada küçük bir rol oynadığına işaret etmektedir.
Her ne kadar, gametik DMR'lerin kimliği ve epigenetik modifikasyonları hakkında çok şey bilinse de, gametlerde metilasyon için bu dizilimlerin nasıl seçildiği konusunda çok daha az bilgi vardır. Şimdiye dek, babasaldan çok daha fazla annesel metillenmiş gametik DMR'ler tanımlanmıştır (Bartolomei & Ferguson-Smith 2011). Annesel metillenmiş DMR'ler, oosit gelişimi sırasında metillenir; babasal metillenmiş DMR'ler ise doğumdan önce prospermatogonia'da metillenir (Fig. 7) (Lucifero et al. 2002). Annesel gametik DMR'ler için, bilinen gametik DMR'lerin dizilim karşılaştırması, herhangi bir çarpıcı dizilim korunumu açığa çıkarmaz ancak bazıları DNA metilasyonu çeken ikincil bir yapı edinmiş olabilen bir dizi doğrudan yineleme içerir (Neumann et al. 1995). Örneğin Igf2r kümesi gametik DMR'deki peşpeşe doğrudan yinelemelerin oosite-özgü DNA metilasyonu için olmazsa olmaz olduğu gösterilmiştir (Koerner et al. 2012). Kcnq1 kümesi gametik DMR'dekiler ise olmazsa olmaz değildir (Mancini-DiNardo et al. 2006). Annesel DMR'lerin bir diğer özelliği de genomun geri kalanına kıyasla kayda değer ölçüde CpG zengini olmalarıdır. Bu bölgelerin nasıl fark edileceği konusunda bir fikir, DNMT3A ve DNMT3L komplekslenmiş karboksi-terminal bölgelerinin X-ışın kristalografisi ile elde edilen yapısal çözümlemesinden gelmiştir (Jia et al. 2007). Bu iki enzimden oluşan bir tetramerik kompleks, tercihan 8-10 baz çifti ayrık olan bir çift CpG çiftini metiller (Cheng 2014). Böyle bir aralık annesel metillemede bulunmuştur ama babasal metillenmiş damgalanmış lokuslarda bulunmamıştır. Ancak bu CpG aralığı, genomda yaygındır; o yüzden böyle bir mekanizmanın özgünlüğünde soru işareti yaratır veya ek özelliklerin gerekli olduğuna işaret eder (Ferguson-Smith & Greally 2007). Ek özgünlük, H3K4 rezidü metillenmemiş ise DNMT3L'nin histon H3'ün amino terminusu ile etkileştiğinin ve yerel DNA metilasyonu başlattığının gösterilmesiyle ileri sürülmüştür (daha fazla ayrıntı için bkz. Ooi et al. 2007; Cheng 2014). Oositteki DMR'lerde de novo DNA metilasyonunun özgünlüğüne katkı yapan bir diğer etken, farklı metillenmiş bölgeler boyunca yazımdır (Chotalia et al. 2009). En önemlisi, sadece, germ hattı ICE'lerini kaplayan protein-kodlayan yazmların DNA metilasyonunun ortaya konmasıyla ilgili olduğu düşünülmektedir. Her ne kadar şu anda bu yazımın DNA metilasyon mekanizmasını nasıl çekebileceği net olmasa da, ICE'ler boyunca yazımın, DNA metilasyonunun oluşturulmasına olanak tanıyan açık kromatin bölgeleri oluşturmak veya sürdürmek için gerektiği ileri sürülmüştür. Mekanizmayı daha fazla incelemek ve tanımlamak için yazım ile de novo DNA metilasyonu arasındaki zamansal ilişkiyi daha ayrıntılı biçimde tanımlamak gerekecektir. Yine de, CpG aralığı, yazım-sonrası histon modifikasyonları ve oositlerdeki yazım, anneye-özgü DNA metilasyon damgalarının edinilmesi için bir başlangıç noktası sağlayabilir.
 Figür-7. Fare gelişiminde genomik damgaları oluştırma, sürdürme ve silme. Germ hattında, ilksel germ hücreleri(PGC) kromatin yapısında çok sayoda değişim geçirir ve jenital yükseltiye (gonad) göç sırasında DNA metilasyonu geçirir. Sonra damgalar cinsiyete-özgü şekilde soy hattında edinilir (yeşil). DNA metilasyonu spesifik olarak babasal ve annesel DNA-metillemeli ICE'lere hedeflenir -doğumdan önce prospermatogoniada ve doğumdan sonra oosit olgunlaşması sırasında. Döllenmeden sonraki global DNA metilasyon değişikliklerine rağmen bu damgalar sürdürülür (turuncu): Zigottaki babasal genomun aktif demetilasyonu ve preimplantasyon embriyodaki pasif annesel demetilasyon. Metilasyonun korunduğu bölge adayları arasında ZFP57 ve PGC7/STELLA sayılabilir. Genomun de novo DNA metilasyonu morula evresinde başlar, bu süre boyunca damgalanmış genlerin metillenmemiş allelleri korunmalıdır. Organizmanın yaşamı boyunca bu damgalar somatik hücrelerde sürdürülürken, ekstra-embriyonik dokulardaki damgalamanın DNA metilasyonunun sürdürülmesine daha az bağlı olduğu düşünülür. Germ hattında, damgalar silinir ve bir sonraki kuşka için yeniden ayarlanır (kırmızı). PTM, yazım-sonrası modifikasyon; MAT, annesel genom; PAT, babasal genom.
Figür-7. Fare gelişiminde genomik damgaları oluştırma, sürdürme ve silme. Germ hattında, ilksel germ hücreleri(PGC) kromatin yapısında çok sayoda değişim geçirir ve jenital yükseltiye (gonad) göç sırasında DNA metilasyonu geçirir. Sonra damgalar cinsiyete-özgü şekilde soy hattında edinilir (yeşil). DNA metilasyonu spesifik olarak babasal ve annesel DNA-metillemeli ICE'lere hedeflenir -doğumdan önce prospermatogoniada ve doğumdan sonra oosit olgunlaşması sırasında. Döllenmeden sonraki global DNA metilasyon değişikliklerine rağmen bu damgalar sürdürülür (turuncu): Zigottaki babasal genomun aktif demetilasyonu ve preimplantasyon embriyodaki pasif annesel demetilasyon. Metilasyonun korunduğu bölge adayları arasında ZFP57 ve PGC7/STELLA sayılabilir. Genomun de novo DNA metilasyonu morula evresinde başlar, bu süre boyunca damgalanmış genlerin metillenmemiş allelleri korunmalıdır. Organizmanın yaşamı boyunca bu damgalar somatik hücrelerde sürdürülürken, ekstra-embriyonik dokulardaki damgalamanın DNA metilasyonunun sürdürülmesine daha az bağlı olduğu düşünülür. Germ hattında, damgalar silinir ve bir sonraki kuşka için yeniden ayarlanır (kırmızı). PTM, yazım-sonrası modifikasyon; MAT, annesel genom; PAT, babasal genom.Eril germ hattında babaya-özgü DNA metilasyon damgalarının nasıl oluşturulduğuna ilişkin çok daha az bilgi vardır. Yine de, ilk deneyler, dişil germ hattı ile bazı benzerlikler olabileceğine işaret etmektedir. Yakın zamanda, damga oluşumu sırasında, ilksel germ hücrelerindeki iki babasal gametik DMR'de (H19-DMD & IG-DMR), baskın olarak tek iplikten, yüksek yazımsal komple-okuma (İng. readthrough) saptandığı gösterilmiştir (Henckel et al. 2011). Ayrıca, DNA metilasyonundan korunan annesel gametik DMR'lerin, eril ilksel germ hücrelerinde H3 lizin 4 trimetilasyon (H3K4me3) bakımından zenginleştirildikleri görülmektedir.
Genomik damgalamadaki en gizemli sorulardan biri, damgalanmış genlerdeki DNA metilasyon işaretlerinin, döllenmeden sonra oluşan genom çapındaki yeni baştan programlamadan (preimplantasyon embriyoda oluşan DNA demetilasyonu ve ardından gelen de novo DNA metilasyon dalgası dahil olmak üzere) nasıl kaçtıklarıdır (Fig. 7) (bkz. Morgan et al. 2005; Li & Zhang 2014). Korumayı sağlayanın, cis-etkir dizilimler ile trans-etkir faktörlerin bir kombinasyonu olması olasıdır. Annesel faktörlerden biri olan PGC7/STELLA, H3K9me2 ile etkileşimler yoluyla erken dönem fare embriyosunda DNA metilasyonunu sürdürmede genel bir role sahip görünür (Nakamura et al. 2012). Ancak damgalanmış genler için daha spesifik olabilecek bir faktör ZFP57'dir. Geçici yenidoğumsal diyabet hastalarında tanımlanan Zfp57 mutasyonlarının çok sayıda damgalanmış lokustaki DNA metilasyon kusurlarıyla ilişkili olduğunu çalışmalar göstermiştir (Mackay et al. 2008). Ek olarak, Zfp57 boş fareler embriyonik ölümcüllük ve çoğu lokusta (ama hepsinde değil) damgalama yitimi sergiler (Li et al. 2008). Daha yakın zamanda, ZFP57'nin kofaktör KAP1'e bağlandığı gösterilmiştir; o da sonrasında başka epigenetik düzenleyiciler görevlendirir (Quenneville et al. 2011). Dolayısıyla, ZFP57'nin dizilime ve DNA metilasyonuna bağlı bağlanışı, KAP1'in allelik bağlanışını belirlemek için bir çapa görevi görebilir; KAP1 daha sonra damgalanmış lokuslardaki heterokromatik, susturulmuş allellere SETDB1, HP1, DNMT1, DNMT3A ve DNMT3B gibi başka başlıca baskılayıcı epigenetik düzenleyicileri görevlendirir. Embriyonun erken evrelerinde, damgalanmış lokuslarda DNA metilasyonunu, henüz tanımlanmamış proteinler sürdürmesi de olasıdır.
3.6 Damgalanmış Gen Kümelerinde Tanımlanan İki Çeşit cis-Etkir Susturma
Şu anda, başlıca iki adet cis-etkir susturma mekanizmaları sınıfının çeşitli kümelerdeki damgalamayı yönettiği hipotezi ileri sürülmüştür: Yalıtıcı modeli Igf2 kümesine uygulanabilir; lncRNA-aracılıklı susturma modeli Igf2r ve Kcnq1 kümelerine uygulanabilir. Bütünüyle tanımlanmış olmasa da, Tablo-1'deki kümelerin çoğu, iki modelden birinin özelliklerini kapsar. Igf2 lokusunda yalıtıcı modelinin tanımına yol açan buluş, H19 yazımının başlangıcından 2 kb yukarı yönde ve Igf2'den 80 kb aşağı yönde konumlanan gametik DMR'nin (H19-DMD) silinmesiydi (Fig. 8) (Thorvaldsen et al. 1998). Silindiği zaman, delesyonun annesel veya babasal kalıtılmasından bağımsız olarak, H19 ve Igf2 damgalama yitimi sergileyerek, bu DMR'nin bir ICE olarak tanımlanmasını sağladı. Daha sonra, bu ICE'nin CTCF'ye (beta-globin lokusunda yalıtıcı etkinliğine aracılık ettiği gösterilen bir protein) bağlandığı ve ICE'nin kendisinin bir yalıtıcı görevi gördüğü gösterildi (Bell & Felsenfeld 2000; Hark et al. 2000). Bu bağlamda, bir yalıtıcı, aralarına konduğunda, yükseltici (İng. enhancer) ve başlatıcı etkileşimlerini engelleyen bir eleman olarak tanımlanır. Dolayısıyla, bu lokustaki damgalanmış gen ifadesi için model şöyledir: Annesel allelde, CTCF ICE'ye bağlanır ve üç genin aşağı yönünde konumlanan H19 lncRNA ile paylaşılan yükselticilere Igf2 ile Ins2'nin erişimini engeller. Böylelikle bu H19'a özel yükseltici erişimi sağlar (Fig. 8). Babasal allelde, DNA metilasyonunu eril germ hattında edinerek, CTCF'nin ona bağlanmasını önler. Dolayısıyla, babasal kromozomda, Igf2 ile Ins2 yükselticilerle etkileşir ve bu kromozomdan ifade edilirler. Babasal ICE'de DNA metilasyonunun varlığı, bilinmeyen bir mekanizma tarafından H19 başlatıcısının ikincil metilasyonuna yol açar ve babasal kromozom üzerinde susturulmuş duruma gelir. Yalıtıcı modeli geniş çapta kabul görmesine karşın, yalıtıcının bu lokusta nasıl davrandığı net değildir. En fazla kabul gören görüşlerden biri, kromatin ilmekleri oluşumu yoluyla genleri yalıtmak için CTCF'nin cis'teki DNA molekülleri ile etkileştiğidir (daha fazla ayrıntı için bkz. Dekker & Misteli 2014). Dahası, bu ilmekleri oluşturmak için kohezinin CTCF ile etkileştiği gösterilmiştir (Nativio et al. 2009). Yalıtıcı modelinde CTCF'nin olması, başka damgalanmış genlerde CTCF bağlanma bölgelerinin tanımlanmasına yol açmıştır; Rasgrf1, Grb10 ve Kcnq1ot1 gibi. Bu da yalıtıcı modelinin diğer damgalanmış kümelerde de işliyor olabileceğine işaret eder.
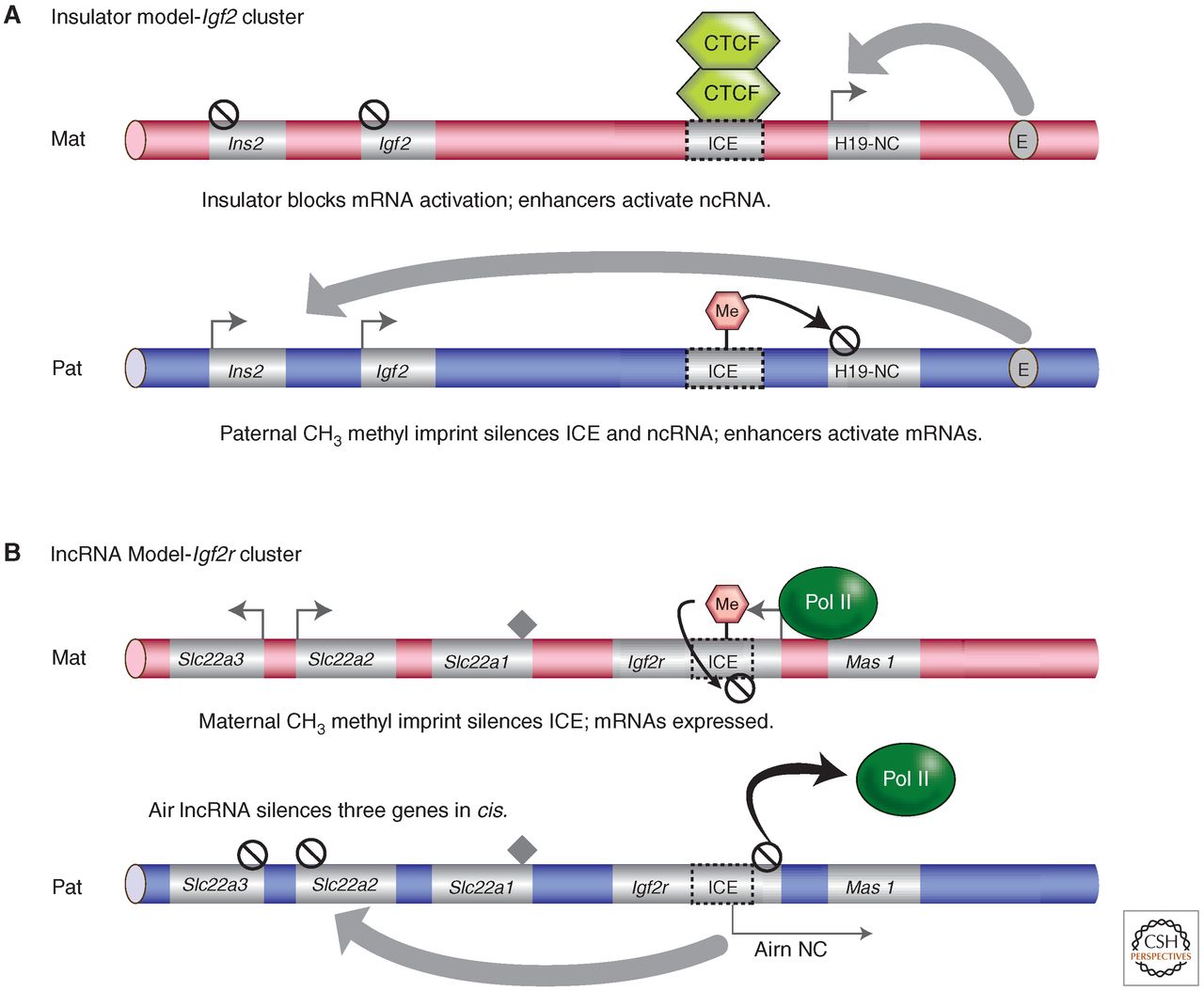 Figür-8. Damgalanmış gen kümelerindeki iki cis-etkir susturma mekanizması. (A) Igf2 kümesi için yalıtıcı modeli. Endoderm için ifade örüntüsü gösterilmiştir. Annesel kromozom üzerinde, metillenmemiş ICE, CTCF proteinine bağlanır ve yaygın endoderm yükselticilerin (E) Igf2 ve Ins2'yi etkinleştirmesini önleyen bir yalıtıcı oluşturur. Yükselticiler onun yerine yakındaki H19 lncRNA başlatıcıyı etkinleştirir. Babasal kromozom üzerinde, metillenmiş ICE, CTCF proteinine bağlanamaz ve bir yalıtıcı oluşturamaz; dolayısıyla Igf2 ve Ins2 mRNA genleri sadece bu kromozom üzerinde ifade edilir. H19 lncRNA metillenir, büyük olasılıkla 2 kb'lık uzaklığındaki metillnemiş ICE'den yayılma nedeniyledir ve susturulur. (B) Igf2r kümesi için lncRNA modeli. Plasenta için ifade örüntüsü gösterilmiştir. Annesel kromozom üzerinde, doğrudan DNA metilasyon damgasıyla susturulmuş olan metillenmiş ICE Airn lncRNA başlatıcı içerir. Igf2r, Slc22a2 ve Slc22a3 mRNA genleri sadece bu kromozom üzerinde ifade edilir. Mas1 ve Slc22a1 plasentada ifade edilmez (boyalı karo). Babasal kromozomda, metillenmemiş ICE'de bulunan Airn lncRNA başlatıcı ifade edilir ve cis'teki Igf2r (kısmen RNA polimeraz II'yi atarak), Slc22a2 ve Slc22a3'ü susturur. Modellerin ikisinde de DNA metilasyon damgasının lncRNA'yı susturduğuna ve mRNA ifadesine izin verdiğine dikkat ediniz. ICE, damga kontrol elemanı; gri ok, damgalanmış genin ifade edilen alleli; çizgili daire, damgalanmış genin baskılanmış alleli; kalın gri oklar, cis'te uzun mesafe etkisi.
Figür-8. Damgalanmış gen kümelerindeki iki cis-etkir susturma mekanizması. (A) Igf2 kümesi için yalıtıcı modeli. Endoderm için ifade örüntüsü gösterilmiştir. Annesel kromozom üzerinde, metillenmemiş ICE, CTCF proteinine bağlanır ve yaygın endoderm yükselticilerin (E) Igf2 ve Ins2'yi etkinleştirmesini önleyen bir yalıtıcı oluşturur. Yükselticiler onun yerine yakındaki H19 lncRNA başlatıcıyı etkinleştirir. Babasal kromozom üzerinde, metillenmiş ICE, CTCF proteinine bağlanamaz ve bir yalıtıcı oluşturamaz; dolayısıyla Igf2 ve Ins2 mRNA genleri sadece bu kromozom üzerinde ifade edilir. H19 lncRNA metillenir, büyük olasılıkla 2 kb'lık uzaklığındaki metillnemiş ICE'den yayılma nedeniyledir ve susturulur. (B) Igf2r kümesi için lncRNA modeli. Plasenta için ifade örüntüsü gösterilmiştir. Annesel kromozom üzerinde, doğrudan DNA metilasyon damgasıyla susturulmuş olan metillenmiş ICE Airn lncRNA başlatıcı içerir. Igf2r, Slc22a2 ve Slc22a3 mRNA genleri sadece bu kromozom üzerinde ifade edilir. Mas1 ve Slc22a1 plasentada ifade edilmez (boyalı karo). Babasal kromozomda, metillenmemiş ICE'de bulunan Airn lncRNA başlatıcı ifade edilir ve cis'teki Igf2r (kısmen RNA polimeraz II'yi atarak), Slc22a2 ve Slc22a3'ü susturur. Modellerin ikisinde de DNA metilasyon damgasının lncRNA'yı susturduğuna ve mRNA ifadesine izin verdiğine dikkat ediniz. ICE, damga kontrol elemanı; gri ok, damgalanmış genin ifade edilen alleli; çizgili daire, damgalanmış genin baskılanmış alleli; kalın gri oklar, cis'te uzun mesafe etkisi.Ancak lncRNA sınıfı damgalama modeli daha yaygın olabilir. Damgalanmış kümelerde işlevsel ncRNA'ların tanımlanmasına yol açan buluş, 108 kb'lık Airn lncRNA'yı 3 kb'a kısaltan bir deneydir (Sleutels et al. 2002). Bu kısaltılmış lncRNA damgalanmış ifadeyi sürdürdü ve Airn başlatıcısı damgalanmış DNA metilasyonunu sürdürmüştür; yine de Igf2r kümesindeki mRNA genlerinin üçünün de susturulması yitirilmiştir (Fig. 8). Ayrıca lncRNA aracılıklı susturmanın Kcnq1 kümesinde işlediği gösterilmiş durumdadır (Mancini-DiNardo et al. 2006); fakat bu dokuya-özgü bir şekildedir ve bu küme (Shin et al. 2008) ile Gnas damgalanmış kümesinde (Williamson et al. 2011), yalıtıcılar kullanan gibi başka bir mekanizmanın da işe karışabileceğine işaret etmiştir. Şu anda, lncRNA'ların genleri nasıl susturduğu tam olarak bilinmemektedir ama çok sayıda model olasıdır. Her bir kümede rastlanan, mRNA ile lncRNA arası sens-antisens çakışmasından iki olasılık doğar. İlk olasılık, mRNA ile lncRNA arasında çift-iplikli RNA'nın oluşması ve RNA-müdahalesi (İng. RNA-interference - RNAi) tetiklemesidir (Martienssen & Moazed 2014). Ancak RNAi düzeneğinin yokluğu, Kcnq1 kümesindeki damgalanmış ifadeyi etkilemez (Redrup et al. 2009). Dolayısıyla, ikinci bir olasılık, bu sens-antisens çakışmasının, yazımı mRNA başlatıcıdan etkileyen bir başlatıcının veya bir yükselticinin bir yazımsal müdahale biçimine neden olmasıdır (Pauler et al. 2012). Bu durumda, ilk önce çakışan başlatıcı veya yükseltici susturulup, sonrasında baskılayıcı kromatin birikiminin yayılması ve küme boyunca yazımsal gen sessizleştirilmesini tetiklemesi gerçekleşebilir. Bu model için kanıt, ES hücrelerdeki (embriyonik kök hücrelerdeki) Igf2r/Airn lokusunda üretilen bir dizi rekombinant endojen (içsel) kromozomlardan gelmiştir (Latos et al. 2012). Embriyoda bu lokustaki allele-özgü ifadenin başlangıcı, Igf2r'nin biallelik olarak ifade edildiği ES hücre farklılaşması tarafından yinelenebilir ama Airn ifadesinin başlangıcı Igf2r damgalaması ile sonuçlanır (Latos et al. 2009). Igf2r susturulması için Airn yazımının mı yoksa lncRNA'nın kendisinin mi gerektiğini sınamak için Airn farklı uzunluklarda kısaltılmıştır; bunun sonucunda susturmanın sadece Airn yazımının Igf2r başlatıcısınınkiyle (RNA polimeraz II görevlendirmesiyle müdahale eder) çalışması gerektiği çıkmıştır (Latos et al. 2012). Bu model, bir lncRNA olarak değil de, Airn'nin baskın olarak yazımı yoluyla etkidiğini ileri sürer. Ancak, X-kromozomu inaktivasyonundaki Xist lncRNA'nın eylemi için tanımlanana benzer bir şekilde, damgalanmış lncRNA'ların yerel kromozomal bölgeyi kaplayarak etkimesi ve doğrudan baskılayıcı kromatin proteinlerini damgalanmış kümeye görevlendirmesi de mümkündür (Brockdorff & Turner 2014: Memelilerde Dozaj Telafisi). Histon yazım-sonrası modifikasyonu mekanizmasının görevlendirilmesinde lncRNA'nın bir işlevine kanıt, plasental dokulardaki deneylerden gelmiştir. RNA floresan in sitı hibritleştirme deneyleri, Airn ve Kcnq1ot1'in, yazım bölgelerinde RNA bulutları oluşturduğunu göstermiştir (Nagano et al. 2008; Pandey et al. 2008; Terranova et al. 2008; Redrup et al. 2009). Terranova ve çalışma arkadaşları, bu uzun ncRNA'ların bir baskılayıcı histon bölmesiyle ve Polycomb grubu proteinlerle ilişkili olduğunu göstermiştir (Terranova et al. 2008). Bu çekirdeksel bölme ayrıca RNA polimeraz II'den yoksundur ve üç boyutlu büzülmüş bir durumda bulunur. Airn lncRNA üzerinde yapılan diğer çalışmalar, lncRNA'ların baskılayıcı histon modifikasyonlarını aktif biçimde görevlendirdiğini söyleyecek kadar ileri gitmiştir (Nagano et al. 2008); ama sadece plasentada. Bu ikinci durumda, Airn'ın EHMT2 H3K9 metiyltransferazı aktif biçimde görevlendirdiği gösterilmiştir. Bu, Igf2r geninin değil ama Slc22a3 geninin babaya-özgü susturulmasıyla sonuçlanır. Bu deneyler, damgalanmış genlerin lncRNA-aracılıklı susturulmasının, farklı aşağı yön mekanizmalarına bağlı olabileceğine işaret eder.
En önemlisi şudur ki, başka damgalanmış gen düzenleme mekanizmaları da muhtemelen vardır. Örneğin, Wood ve çalışma arkadaşları, allele-özgü bir şekilde alternatif poliadenilasyon bölgelerinin kullanıldığı yeni bir damgalanmış lokus (H13) tanımlamıştır (Wood et al. 2008). H13 geni, DNA metilasyonunu oositlerde edinen annesel metillenmiş bir içsel (İng. internal) CpG adası içerir (henüz ICE aktivitesi bakımından test edilmemiştir). Bu CpG adasının hipermetilasyonu, annesel kromozomdan tam uzunluklu ve işlevsel H13 gen yazımının sentezini garantiler. Babasal allel üzerindeki metillenmemiş CpG adasının, Mcts2 retrogeninin başlatıcısından yazıma izin verdiğini deneyler göstermiştir. Bunun karşılığında Mcts2 ifadesi, H13'ün prematüre poliadenilasyonuyla ve dolayısıyla kısaltılmış H13 yazımlarının ifadesiyle bağlaşıktır. Bu lokus, damgalanmış genlerin eksiksiz bir kataloğu ortaya konduğunda, daha ender kullanılan diğer genomik damgalama mekanizmalarının tanımlanma olasılığını yükseltmiştir.
4. Genomik Damgalama: Memeli Epigenetik Düzenlemesi İçin Bir Model
Genomik damgalamayı incelemek, diğer memeli epigenetik gen düzenleme modellerinden daha avantajladır çünkü hem aktif hem de inaktif ebeveynsel allel aynı çekirdekte bulunur ve aynı yazımsal ortama maruz kalır (Bartolomei 2009; Barlow 2011). Bunun sonucunda, epigenetik değişikliklerin hücrenin değişmiş farklılaşma durumunu da yansıtabildiği "önce ve sonra" epigenetik sistemlerin aksine, iki ebeveynsel allel arasındaki herhangi bir epigenetik farkın yazımsal durumlarıyla bağlaşık olması daha muhtemeldir. Hem aktif hem de sessiz ebeveynsel allelin aynı çekirdekte olması, epigenetik gen düzenlemeyi incelemek için genomik damgalamayı ideal bir sistem yapar. Aynı zamanda, bir güçlük çıkarır çünkü önce ebeveynsel allelleri ayırt etmek gerekir ki böylece gen etkinliği ve sessizleşmesi ile ilişkilendirilen spesifik özellikler, doğru ebeveynsel allele atfedilebilsin. Farede, annesel ve babasal kromozomların ayırt edilmesini sağlayan model sistemlerin geliştirilmesiyle, bu güçlüğün büyük ölçüde üstesinden gelinmiştir (Fig. 1). Epigenetik gen düzenleme mekanizmalarının evrimde yüksek ölçüde korunduğu gerçeğine karşın, her bir organizma için genom organizasyonunun çeşidi ile ilişkili farklılıklar olması muhtemeldir. Memeli genomu, yinelemelerin (yeri değişebilen elemanlar olarak da bilinir) çok sayıda kopyasının arasına genleri serpiştiren alışılmadık bir organizasyon sergiler. Bu, çoğu genin uzunluğunu epeyce yükseltmesinin yanı sıra, komşu genler arası uzaklığı da yükseltir. Bu, genomları yinelemesiz kalma ya da hiç değilse genleri yinelemelerden ayırma yönünde bir eğilim sergileyen maya, nematod, bitki ve Drosophila gibi diğer model organizmalarla tezat oluşturur (organizmaların karşılaştırması için bkz. Rabinowicz et al. 2003; Allis et al. 2014'teki Fig. 19). Memeli epigenetiğinin kavranmasında genomik damgalama nasıl katkıda bulunabilir? Damgalanmış gen kümelerinin karakterizasyonu tamamlanmaktan uzak olmasına rağmen, yerel bölgelerde ve alanlarda genlerin nasıl kontrol edildiği hakkında bilgi sağlama potansiyellerinin olduğu açıktır. Şu âna dek, damgalanmış gen kümeleri hâlihazırda şunlara örnekler sağlamış durumdadır: DNA metilsyonu tarafından düzenlenen (regüle edilen) cis-etkili DNA dizilimleri, memeli genomunda kendiliğinden sessiz olan ve ifade edilmeleri için epigenetik etkinleştirme gereken genler, yalıtıcı görevi görebilen uzun-erimli düzenleyici elemanlar ve cis'te büyük gen alanlarını sessizleştiren sıradışı lncRNA'lar. Bu epigenetik düzenleme mekanizması çeşitlerinin damgalanmış kümelere mi özgü olduğunu, yoksa memeli genomundaki damgalanmamış genlerin ifadesinin düzenlenmesinde de mi bulunabildiklerini zaman gösterecektir.
5. Geleceğe Yönelik Düşünceler
1991'de memelilerdeki ilk damgalanmış genlerin keşfinden bu yana, genomik damgalama yoğun bir ilginin odağında bulunmaktadır. Eski deneyler damgalanmış genleri belirlemek için moleküler ve genetik stratejilere dayanırken, polimorfik bireyler üzerindeki yüksek-çıktılı teknoloji damgalanmış genlerin (Deveale et al. 2012) ve ebeveyne-özgü DNA metilasyonu içeren bölgelerin (Xie et al. 2012) eksiksiz belirlenmesine olanak tanımaktadır. Bu deneyler, yaygın damgalanmış ifade sergileyen genlerin çoğunun zaten tanımlanmış olduğuna işaret etmektedir (http://www.mousebook.org/catalog.php?catalog=imprinting). Bununla birlikte, dokuya-özgü damgalanmış ifade sergileyen bazı genlerin henüz tanımlanmamış olması olasıdır (Prickett & Oakey 2012). Bazı sorular hâlâ nihai yanıtlarını beklemektedir; özellikle de omurgalılar arasında neden sadece memelilerin embriyonik ve yenidoğansal gelişimi düzenlemek için damgalanmış genleri kullandıklarıyla ilgili olanlar. Bu bilgi yoksunluğu, memelilerde damgalanmış ifadey, kontrol eden epigenetik mekanizmaların gün yüzüne çıkarılmasıyla geçen 20 yıl boyunca süren kapsamlı ilerlemeyle tezat oluşturur. Her ne kadar tüm ayrıntılar hâlâ netleşmiş olmasa da, bu bilgi sayesinde, damgalanmış gen kümelerinde damgalama mekanizmasının işleyişinin genel ilkelerini anladığımızı düşünüyoruz. Şu aşamada, ebeveyne-özgü ifade düzenlemek için genomik damgalamanın hücrenin normal epigenetik mekanizmasını kullandığı ve bu mekanizmanın gamette sadece bir ebeveynsel allele sınırlandırılmasıyla her şeyin harekete geçtiği açıktır. Farklı gen kümelerinde damgalanmış ifadeyi kontrol eden mekanizmada genel benzerlikler olmasına karşın, memeli genomunda bu mekanizmanın kaç tane varyantının olduğu henüz bilinmemektedir. Damgalanmamış genlerin hangi dereceye kadar damgalanmış gen kümeleri için tanımlanmış epigenetik mekanizmalar tarafından kontrol edildiklerini belirlemek, ileride ilgi çekici olabilir. Nihayetinde, bu bilginin insanlarda tedavi amaçlı kullanılması, örneğin Prader-Willi ve Angelman sendromu hastalarında sessiz ebeveynsel allellerin yeni baştan ifadesinin tetiklenmesiyle, semptomlarının hafifletilmesinde yararlanılabilir (bkz. Huang et al. 2011; Beaudet & Zoghbi 2014). Epigenetik düzenlemenin kanserlerde (Jones & Baylin 2014), yardımlı üreme teknolojilerinde ve ayrıca yaşlanma sürecinde (Rando & Chang 2012; Berger & Sassone-Corsi 2014) dengesizleşebileceğinin fark edilmesiyle birlikte, hücrenin epigenetik bilgiyi kontrol etme yolunun anlaşılmasının önemi giderek artıyor. Genomik damgalamanın daha iyi bir kavrayışının, memeli genomunun gen ifadesini düzenlemek için epigenetik mekanizmaları nasıl kullandığının keşfedilmesi için önemli bir model olmayı sürdüreceğine kuşku yok.
- Genomic Imprinting in Mammals https://cshperspectives.cshlp.org/content/6/2/a018382.full
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek OlBunlar da İlginizi Çekebilir








En Çok Okunan
Bu Ay Öne Çıkanlar



İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.