
 Petrovich9 / iStockPhoto
Petrovich9 / iStockPhoto

Gezegenimizdeki canlılara baktığımızda çok sayıda farklı üreme yöntemi görüyoruz. Fakat hepsini temel olarak iki ana başlık altında inceleyebiliriz: Eşeysiz üreme ve eşeyli üreme. Canlı türlerinin büyük bölümü bu iki üreme türünden sadece biri ile çoğalabilirken, bazıları ise hem eşeyli hem de eşeysiz üreme becerisine sahip olup, farklı koşullara bağlı olarak iki şekilde de çoğalabiliyor.
Sadece eşeysiz üreyen türlerde herhangi bir cinsiyet söz konusu değil. Seçimli veya zorunlu olarak eşeyli üreyen türlerde ise cinsiyet özelliği mevcut. Bu özelliği daha iyi anlayabilmek için eşeyli üremede kullanılacak üreme hücrelerinin oluşumunu sağlayan “mayoz bölünme”yi ve eşey(cinsiyet) kavramını inceleyelim.
Mikroplarda Cinsiyet
Mikro ölçekteki canlılar arasında eşeysiz üreme yaygın bir yöntem olsa da, eşeyli üreyen mikroplar da vardır. Ökaryot mikroplar, bakterilerden farklı olarak ikiye bölünmeyle çoğalmaz; hemen hepsi eşeyli ürer. Bitkiler ve hayvanlar gibi büyük ölçekli ökaryotlarda cinsiyetlerin farkına varmak kolaydır ama mikroplarda bu nasıl anlaşılabilir? Doğrudan olmasa da, çeşitli gözlemlerle dolaylı yoldan bilgi edinilebilir: Homolog rekombinasyon olması, sitoplazmik kaynaşma olması, karmaşık yaşam çevrimlerinin doğrulanması veya potansiyel üreme hücrelerinin üretimi gibi. Ayrıca genetik açıdan eşeyli üreme ile ilişkilendirilen iki tür genin var olup olmadığı kontrol edilerek de fikir edinilebilir. Bu genler Eşleşme Tipi Lokusu (MAT lokusu) ve mayoza özgü genlerdir.
Eşleşme Tipi Lokusu olan MAT lokusu çok sayıda ökaryotik mikrop türünde keşfedilmiştir ve eşleşme tipinin belirlenmesini kontrol eder. Cryptococcus neoformans türü canlıda MAT lokusu 2 farklı eşleşme tipi ifade eder: a ve alfa. Tetrahymena thermophila türünde ise iki allelin programlanmış genom düzenlenmesiyle etkileşimi yoluyla 7 farklı eşleşme tipi kodlar. Physarum polycephalum’da 13 tane, basidiomycetes’te binlerce eşleşme tipi vardır. Çeşitli canlılarda daha farklı eşleşme tipi sayılarına da rastlanmıştır. Bir mikrobun genomunda MAT lokusunun var olması, mikrobun eşeyli üreme olasılığını temsil eder.
Mayozla ilişkilendirilen proteinleri kodladıkları bilinen genlerin bir genomda bulunması da, canlının eşeyli ürüyor olma olasılığını yükseltir. Fakat örneğin Giardia genomunda mayoza özgü genlerin 7 tane homologu olmasına rağmen, bu canlıda mayoz hiç görülmemiştir. Organizmanın daha karmaşık bir yaşam çevrimi vardır ve bu genlerin varlığı, organizmada genetik varyasyonun nasıl kontrol edildiğini belirlemek amacıyla kullanılır. Her ne kadar organizmada mayoz gerçekleşmiyor olsa da, homologların varlığı, benzer bir kontrolün olabileceğine veya bu cinsin son ortak atasının yaşam çevriminde mayozun kullanıldığına işaret eder.
Cinsiyeti Olan İlk Canlı
Eşeyli üreme, ökaryotik organizmaların neredeyse tümünde var olan bir özellik olduğundan, tüm ökaryotların son ortak atasında (Son Ökaryotik Ortak Ata – SÖOA ) ortaya çıktığı düşünülüyor. Prokaryottan proto-ökaryota ve ondan da ökaryota doğru ilerleyen geçiş sürecinde mutasyon, yatay gen aktarımı, endosimbiyoz ve seçilim yoluyla, önceden var olan genetik devrelerin korunumu, üzerlerinde değişiklik yapılması ve yeni düzenlemeleri belli bir çerçevede gerçekleşti. Bu evrimsel süreçte, haploid gametlerin düzenlenmiş kaynaşmalarını ve diploidden haploide mayoz yoluyla indirgemeleri kapsayan işlemler sonucunda kromozomların yeniden düzenlenmesi ve farklı şekilde bir araya getirilmesi (rekombinasyon) ile SÖOA eşeysel özellik kazandı. Cinsiyetin ve mayoza özgü genlerin belli başlı tüm ökaryotik dallarda keşfedilmesi dolayısıyla, SÖOA’nın eşeysel özelliğe sahip olduğu konusunda bilimciler görüş birliğine varmış bulunuyor. Şu anda eşeysiz üreyen ökaryotik mikropların genomunda MAT lokusunun var olması da, onların evrimsel süreçte eşeyli üreme özelliklerini yitirdiklerine işaret ediyor.
Cinsiyetsizlikten cinsiyetliliğe geçiş, dört temel yenilik gerektiriyor:
- Hücre-hücre kaynaşması ve mayoz yoluyla ploidinin (homolog kromozom kümesi sayısı) değişimi,
- hücre-hücre kaynaşmalarında eşleşme tiplerinin (İng. mating-type), bütünleyici haploid gametlerin (izogametik ve daha sonra anizogametik) farklılaşması yoluyla düzenlenmesi,
- diploid/mayotik durumun eşleşme tipine bağlı çiftlenmesiyle uyumlanabilir diploid uyuyan sporların (İng. resting spore) oluşumu ve
- organel genomlarının eşleşme tipine bağlı aktarımı.
Proto-ökaryotton SÖOA’ya geçiş döneminde, cinsiyet açısından çok sayıda deneysel örneğin ortaya çıkmış olması muhtemeldir. Bunların büyük bölümü soy sürdürmeyi başaramazken, bir kısmı ise eklemlenmiş, bütünleşmiş ve üzerlerinde değişiklikler gerçekleşmiş olabilir. Dolayısıyla yukarıdaki dört maddelik listeyi bir olay dizisi olarak değil de, birbirlerine paralel olarak birlikte evrilmiş yenilikler olarak düşünmek daha uygundur. Bu eşeysel çevrim özelliklerinin ortaya çıkmasıyla birlikte, ökaryotik cinsiyetin evrimi sayısız farklı pre-zigotik ve post-zigotik varyasyona olanak sağlamış olabilir. Bunun sonucunda da panmiktik popülasyonlar, farklı uyumlanmaları olan ayrık türlere bölünmüştür.
Hücre-Hücre Kaynaşması
Bakterilerin dış kaynaklardan DNA alabilmek gibi bir becerileri vardır. Muhtemelen proto-ökaryotlar da evrimlerinin erken evrelerinde fagositik davranışa sahipti; en azından arada-sırada. Ne de olsa elimizde mitokondri gibi sağlam bir kanıt var: Bir zamanlar bir yerde bir proto-ökaryot, yabani bir proteobakteri yakalayıp evcilleştirerek, onu organel hâline getirdi. Modern hücrelerde fagositoz çok karmaşık bir süreçtir ve yüzlerce protein rol alır. Ama erken dönem versiyonlarında büyük olasılıkla işler daha basitti ve fagositoz olayına girebilen hücre zarları, muhtemelen hücre-hücre kaynaşması yapma becerisine de sahipti; modern bakterilerin duvarsız mutantlarında olduğu gibi.
Erken dönem hücre-hücre kaynaşmalarının, “bencil” transpozonlar ve plazmidler tarafından desteklenmiş olabileceği ileri sürülmüştür; böylece yepyeni genomik koşullarda kopyalanma bakımından avantaj elde edebilirlerdi. Bu hipotezin olası bir kanıtı, bazı mayalarda eşleşme tipleri arası geçiş etkisi yaratmak için atasal “yeri değişebilen elemanlar”dan (İng. transposable elements) alınmış olan bir transposazın veya HO endonükleazın kullanımıdır.
Hücre-hücre kaynaşması, kromozom sayısında bir artış yaratır. Modern ökaryotik genomların geniş büyüklükleri endomitozun sonucu olabileceği gibi, proto-ökaryot evrimi sırasında genom genişlemesine hücre-hücre kaynaşmalarının katkı yapmış olması da olasıdır. Aynı sonuca götüren alternatif bir olasılık da, hücre-hücre kaynaşmalarının ilk örneklerinin bir proto-ökaryotun bir diğeri tarafından kapsanması şeklinde olması ve içte kalan hücre zarının ya sindirilmiş ya da dış hücrenin zarıyla içerden birleşmiş olmasıdır.
Kromozom sayısının artışının, yeni fırsatlara olanak tanıyacağı kuşkusuzdur. Daha önce sahip olmadığı genler edinmesi, organizmaya bulunduğu koşullarda işine yarayabilecek beceriler geliştirme şansı yaratır. Hızlı ploidi geçişlerinin organizmanın yararına olabilmesi de, eşeyli üremenin rekombinasyondan ayrı bir iyi yanıdır. Haploidden diploide ve sonra tekrar diploidden haploide geçiş yapma becerisi ise ploidi indirgeyici bir mekanizma gerektirir. Modern ökaryotlarda bu işlem mayotik veya paraseksüel süreçlerle gerçekleşir.
Paraseksüellik
Hücrelerin kaynaşması öncelikle ilk paraseksüel çevrimlerin ortaya çıkmasına yol açmış olmalıdır. Daha sonra bu çevrimlerden, gerçek mayotik cinsel çevrimler evrilmiş olabilir. Alternatif bir görüşe göre ise mayoz, öncesinde herhangi bir paraseksüel deneme olmadan, diploid atadan haploid döl üretmenin bir yolu olarak erken bir zamanda doğmuş olabilir. İlk mayozlar muhtemelen düzensiz ve hatalı gerçekleşiyordu; belki ploidinin paraseksüel değişiminden birazcık daha iyiydi. Zamanla daha hatasız mekanizmalar evrilmiş olabilir.
Paraseksüellik gibi mayoz da zararlı mutasyonlardan kurtulmayı sağlar. Ayrıca heterozigot bir diploid atanın haploid yavrusundaki çekinik allellerin açığa çıkmasına olanak tanır. Diploid duruma geri dönüş ilk başta endoreplikasyon ile gerçekleşmiş ve hücre-hücre kaynaşması mekanizmaları daha sonra evrilmiş olabilir. Paraseksüel mekanizmalara kıyasla mayozun avantajları hatasızlık (İng. fidelity) ve daha geniş bir kromozomlar-arası rekombinasyondur; en azından bugün var olan mayoz biçimleri için durum böyledir.
Özetle mayoz şu dört işlevi yerine getirir: (1) Ploidiyi indirger (bunu paraseksüellik de yapar ama daha verimsiz olarak); (2) zararlı allellerden kurtulmayı ve mayotik ürünler haploid olduğunda yararlı çekiniklerin açığa çıkmasını sağlar (paraseksüellik bunu da yapar); (3) ploidiyi indirgerken, eksik ya da düzensiz olmayan tam kromozom kümeleri üretir (mayoz I’in zerafeti); (4) heterozigotlarda krosover ve bağımsız toplama yoluyla rekombinant döl üretir. Paraseksüelliğin ve mayoz temelli cinsel çevrimlerin kökenleri ve evrimi üzerinde düşünürken, eşleşme tipi temelli cinsel farklılaşmaları hâlihazırda ortaya konmuş olan iki modern organizma üzerinden gidelim.
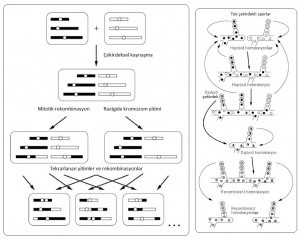
Sağda: Filamentli Ascomycetes’de paraseksüel çevrim.
Hücresel kaynaşma, çekirdeksel kaynaşma ve haploidleşme sonucunda,
üstteki ebeveynlerden alttaki yavrular oluşur.
(1) Aspergillus’ta Paraseksüellik Aspergillus aslında homotallik (zigot oluşturmak için kaynaşabilen iki tür gamet üreten haploid bir evresi olma durumu; genellikle alglerde ve mantargillerde rastlanan bir özelliktir) bir cinsel çevrime sahiptir ama laboratuar koşulları altında paraseksüel çevrime de girebilir: Haploid atalardan, hifal kaynaşma (İng. hyphal fusion) ile diploidler üretilir ve ardından ortaya çıkan diploid rastgele kromozom yitirerek, haploid duruma geri döner. Yakın zamanda yapılan çalışmalarda, bu paraseksüel çevrimin, evrim için kapasitör görevi görebilecek korunaklı bir diploid duruma giriş sağlayabileceği gösterilmiştir.
Daha hızlı büyüyen varyantlar, homozigot diploidlerden kolayca ortaya çıkar (ama karşılık gelen haploid “ebeveyn”den çıkmaz) ve ardından haploid duruma indirgenirler. Hızlı büyüyen varyantların çok sayıda çekinik allel taşıdığı görülmüştür. Bunlar karşılıklı olarak epistatik (birbirlerini baskılayıcı) olup, birlikte yararlı fakat tekil olarak zararlıdır. Dolayısıyla sadece diploide birikebilirler ve daha sonra paraseksüel genom indirgenmesinde salınırlar. Bu çalışmalar sonucunda, haploid-diploid-haploid paraseksüel çevrimlerin, bütünüyle yeni genotipik ve fenotipik çeşitlilik üretme kapasiteleri anlaşılmıştır.
(2) Candida albicans’ta Paraseksüellik Mantarsı bir insan patojeni olan Candida albicans’ın sıra dışı bir paraseksüel çevrimi vardır. Eşleşme tipi lokusu homozigot olduğunda ve hücreler “opak” adı verilen özelleşmiş bir üreme hücresi tipine dönüştüğünde, diploidler arasında çiftleşme gerçekleşir. Böylece hücre-hücre kaynaşması tetraploid hücreler üretir ve olumsuz ortam koşullarının tetiklemesiyle gerçekleşen rastgele kromozom yitimi sonucunda, paraseksüel süreç yoluyla diploid veya diploide yakın duruma dönülür. Candida’nın eşeyli mantargillerden biri olduğu düşünülürse, bu muhtemelen sonradan edinilmiş bir paraseksüel durumdur. Fakat paraseksüel ploidi değişiminin takip ettiği hücre-hücre kaynaşmasıyla genetik değiş-tokuşun buna benzer eski versiyonlarını gözümüzde canlandırabiliriz. Gerçek seksüel çevrimler de mayozun icadıyla onlardan evrilmiştir. Bu açıdan bakıldığında, mayozun evrimi, cinsel çevrimlerin evriminin ilerleyen dönemlerinde gerçekleşmiş olabilir.
C. albicans’ın paraseksüel çevriminde, kromozom kaybına ve toplanmasına ek olarak, düşük düzeyli bir kromozomal rekombinasyon da saptanmıştır. Bu rekombinasyon olayları, mayozda merkezi rol oynayan C.albicans Spo11 ortologunun işlevini gerektirir. Bir yoruma göre, Spo11’in işlevleri mitotik, paraseksüel bir rol oynamak için yeniden düzenlenmiştir. Alternatif olarak, C.albicans’ın paraseksüel çevrimi mayozun bazı özelliklerini (örneğin Spo11’e bağlı kiyazmata gibi) içeriyor olabilir ama ortaya çıkan anöplodinin oranının yüksekliğine bakılırsa, bu doğru sonuçlar vermez ve “paramayoz” gibi bir şey olarak düşünülebilir. Başka mayotik ortologların rol oynayıp oynamadığının incelenmesi, atasal bir paraseksüel-eşeysel geçiş aşamasına benzeyen edinilmiş bir durumu temsil eden bu ilginç örneği aydınlatabilir.
Çok sayıda çalışma, sadece mayoza giren hücrelerde ifade edilen bazı genler belirlemiş ve böylece mayozun alet çantasındaki gereçler gibi düşünülen “mayoza özgü genler”den söz edilmeye başlanmıştır. C.albicans genomunda ise bu çantadaki gereçlerin hepsinin olmadığı anlaşılmıştır; ki bu da onun paraseksüel çevrimiyle uyumlu görünmektedir. Yakın akrabalarında neler olup olmadığını görmek için paraseksüel, aseksüel ve tam seksüel olan çeşitli Candida türlerinin genom dizilimleri çıkarılmıştır. Dikkate değer biçimde, çok önemli olan eşleşme tipi lokusu genlerinin, seksüel olan iki türde eksik olduğu ama yine de cinsel açıdan doğurgan oldukları keşfedilmiştir. Dahası, diğer iki düzine türle beraber seksüel türlerde de, C.albicans’da olmayan önemli mayotik genlerin aynılarının olmadığı görülmüştür. Candida lusitaniae adındaki sıra dışı seksüel türün gerçek anlamda mayoza girmesinin deneysel olarak belgelenmesi dolayısıyla, mayozu gerçekleştirebilmek için gereken gen repertuvarının sanılandan çok daha esnek olduğu ortaya çıkmıştır. İlginç bir şekilde bu eşeyli tür yaklaşık olarak üçte iki oranında öploid döl üretirken, üçte bir oranında anöploid veya diploid verir. Bunun nedeni muhtemelen önemli bileşenlerin eksik olmasından ötürü, mayozu düzgün gerçekleştiremeyişidir. Bir görüşe göre, mayozun tam bir doğrulukla gerçekleştirilmeyişi (İng. meiotic infidelity), bir başka çeşitlilik kaynağı olarak da görülebilir.
Mayoz İçin Gereken Altyapı
Mayozun paraseksüel çevrimlerden evrildiği görüşünde de, öncesinde paraseksüel deneme olmadan erken tarihte evrildiği görüşünde de, mayoz için gereken enzimler ve mekanizmanın, hem bakteri soylarından hem de arke soylarından gelen prokaryotik atalardan alınıp, gereksinimlere göre modifiye edilmiş temel bir “DNA-yönetme enzimleri kümesi”nden evrildiği var sayılır.
Örneğin DNA çift-iplik krılmaları sağlayarak mayotik rekombinasyona yol açan Spo11, proto-ökaryot genomuna dahil edilen atasal arkesel bir topoizomeraz VI homologtan türetilmiştir ve daha sonra mayoza uyumlu hâle getirilmiştir. Kısacası Spo11, DNA’yı yeniden bağlama becerisini yitirmiş bir topoizomerazdır. Şimdiye dek incelenen tüm belli başlı ökaryotik soylarda var olması dolayısıyla, SÖOA’da Spo11’in var olduğu çıkarımı yapılabilir. Bunun dikkat çekici istisnaları ise Dictyostelium discoidium ile yakın akrabası olan cıvık mantarlardır. Bunlar görünüşe bakılırsa Spo11’lerini yitirmiştir ama yine de eksiksiz mayotik cinsel çevrimlere girerler.
Çizgisel Kromozomun Ortaya Çıkışı
Mayozda kromozom ayrışması olur. Bu nedenle kromozomların yapısı üzerinde düşünmek önemlidir. Modern prokaryotların çoğu halka biçiminde genomlara sahiptir. Proto-ökaryotların çizgisel kromozomlarının olduğunu ya da en azından bunların mayoz evrildiğinde ortaya çıktığını öne sürmek için çeşitli sebepler vardır. İlk proto-ökaryotlar muhtemelen bir ya da birkaç tane çizgisel kromozoma sahipti (düzgün ayrışma ve uçların bozunumdan korunmasını sağlayacak olan sentromerler ile telomerlerin evrilmesi için de bu gereklidir). Çizgisel kromozomlar gerekliydi çünkü iki dairesel kromozom arasındaki krosover, kararsız olan disentrik bir halka üretir (eğer iki sentromer karşıt uçlu mikrotübüllere bağlanırsa kromozom kırılır). Bir çizgisel kromozom ile bir dairesel kromozom arasındaki krosover bile kararsız bir sonuç verir.
Eşeysel çaprazlanmaların ortaya çıkışı, pekâlâ çizgisel kromozomlara yol açmış olabilir. Atasal bir proto-ökaryot, sentromerleri olan iki dairesel kromozoma sahip olmuş ve bunların rekombinasyonu dairesel bir disentrik ortaya çıkarmış olabilir. Eksik hâldeki kırılma-kaynaşma-yeniden birleşme çevrimlerine maruz kalınca, çizgisel kromozomlar ortaya çıkmış olabilir. Alternatif olarak, bazı bakterilerde ve bazı organel genomlarında olduğu gibi çizgisel kromozomlar başka şekillerde ortaya çıkmış da olabilir ve eşeysel çaprazlanmalardan daha eskiye dayanıyor olmaları olasılığı da vardır.
Mayozun Evrimi Hakkındaki Tarihsel Tartışmalar
Son derece karmaşık sitolojik özelliklere ve genetik sonuçlara sahip bir hücresel süreç olan mayoz, çok hücreli ökaryotlarda genelde şunları içerir:
- İki çekirdeksel bölünme,
- Kromozom tanıma,
- Kromozom çiftlenmesi,
- Genetik rekombinasyonlu krosover,
- Kromozom sayısının indirgenmesi ve dolaylı (çünkü hücresel ve çekirdeksel kaynaşmanın kromozom sayılarının indirgenmesinden önce gerçekleşmesi gerekir) olarak
- Hücresel tanıma,
- Hücre kaynaşması
- Çekirdeksel kaynaşma.
İlk bakışta bu kısmi süreçlerin (örneğin kromozom çiftlenmesinin ya da hücre kaynaşmasının) hiçbiri, diğer kısmi süreçler olmadan seçimsel avantaj yaratamayacak gibi görünür. Darlington 1958 yılında bu problem karşısında şunları söylemiştir: “O halde mayozun ve eşeyli üremenin kökeni, tüm evrim sürecindeki en şiddetli süreksizliği temsil etmelidir. Ani bir değişimin de ötesinde, bir devrime gerek vardır. Bu olayı, her biri adaptasyon değeri taşıyan değişimlerin aşamalı (gradüel) birikimi olarak hayal etmek imkansızdır. Eğer eşeyli üremenin ardındaki maddesel süreçler o zamanlar anlaşılmış olsaydı, ne Lamarck ne de Darwin evrimi tümüyle süreklilik sergileyen varyasyonların uyumlanarak birikimine bağlamayı akıllarından bile geçirmezlerdi.”
Mayozun kökenlerine ilişkin sitolojik olaylar da, seçimsel baskılar sorusu kadar kafa karıştırıcı olmuştur. Mayozun mitozdan evrildiği neredeyse kesin olmakla birlikte, ilk bakışta dört yeni basamak göze çarpar:
- Homolog kromozomların çiftlenmesi,
- Çiftlenme sırasında kardeş olmayan kromatitler arasında geniş ölçekli rekombinasyon olması,
- İlk mayotik bölünme sırasında kardeş kromatitlerin ayrılmasının baskılanması ve
- İkinci mayotik bölünme sırasında kromozom replikasyonunun olmaması.
Bu özelliklerin hepsinin tek aşamada eşzamanlı olarak belirmesi olanaksız gözükürken (yakın zamanda öne sürülen hipotezlerde mayozun aslında daha kısa ve verimli şekilde gerçekleşmesi kuramsal olarak olanaklıyken, olduğu şekilde olmasının yeniliğe değil, mitozdan türemeye işaret ettiğine dikkat çekilmiştir), haploid kromozom kümelerinin güvenilir biçimde üretimi için tüm dizilimin gerekli olduğu düşünülürse, farklı mutasyonların seçilimi yoluyla adım adım edinilmelerinin de pek kolay olmadığı ortadadır. Tüm bu nedenlere bağlı olarak, mayozun kökeni problemi, Maynard Smith tarafından en zor evrimsel problemlerden biri olarak görülmüştür.
Hamilton ise şunları yazmıştır:
“Evrimsel olarak arka arkaya gelen tüm olaylar arasında, hayretimin kendi zihnim tarafından çözümleme içgüdümü hâlâ yendiği ve Darwinci bir aşamalılık (İng. gradualism) görmekte zorlandığımı itiraf etmem gereken tek bir olay varsa, o da mayozun ortaya çıkması olayıdır.”
Mayoz bölünmenin evrim sürecinde tam olarak hangi aşamalardan geçtiği konusunda henüz görüş birliği sağlanabilmiş değildir. Mayozun mitozdan evrilmiş olduğu noktasında bilimciler hemfikir olsa da, tartışma mayozun evrimini açıklamak için doğal seçilimin yeterli olup olmadığı etrafında dönmektedir. Elde edilen yeni bilgilere dayanarak, konuya ilişkin yanlışlanabilir hipotezlerin geliştirilmesine ve bu hipotezlerin, bilimsel araştırma yöntemleriyle sınanmasına devam edilmektedir. Doğal seçilimin yanı sıra başka hangi mekanizma ya da münferit olayın mayozun ortaya çıkışında rol almış olabileceği konusunun, genç araştırmacıların yapacağı katkılarla aydınlanacağı umuluyor.- What-When-How, "Parasexual Cycle (Molecular Biology)" http://what-when-how.com/molecular-biology/parasexual-cycle-molecular-biology/
- MicrobeWiki, "Sexual Reproduction of Pathogenic Eukaryotes". https://microbewiki.kenyon.edu/index.php/Sexual_Reproduction_of_Pathogenic_Eukaryotes
- Adam S. Wilkins and Robin Holliday, “The Evolution of Meiosis From Mitosis”. GENETICS January 1, 2009 vol. 181 no. 1 3-12 https://doi.org/10.1534/genetics.108.099762
- Ursula Goodenough and Joseph Heitman, “Origins of Eukaryotic Sexual Reproduction”. Cold Spring Harb Perspect Biol 2014
- David Penny, “The evolution of meiosis and sexual reproduction”. Biological Journal of the Linnean Society (1985), 25: 209-220
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek OlBunlar da İlginizi Çekebilir








En Çok Okunan
Bu Ay Öne Çıkanlar



İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.