
Hücreler Neden Proton Derecesine Bağlı Çalışır?
 © 1999 Nature Publishing Group Orgel, L. Are you serious, Dr Mitchell? Nature 402, 17 (1999).
© 1999 Nature Publishing Group Orgel, L. Are you serious, Dr Mitchell? Nature 402, 17 (1999).

Neredeyse tüm hücreler, zarlarının ötesine proton (hidrojen iyonu) pompalayarak "nefes" alır. Moleküler biyolog Leslie Orgel'e göre bu, biyolojide Heisenberg, Schrödinger ve Einstein'ın ortaya attığı sıradışı fizik fikirleri ile kıyaslanabilecek tek düşüncedir. Uçuk İngiliz biyokimyacı Peter Mitchell öncülüğünde ve kendi laboratuvarında geliştirilen bu fikir, 20 yıldan uzun bir süre boyunca tartışmalı olarak kaldı. Bu dönem (solunumdaki ATP sentezi mekanizması olan "oksidatif fosforlanma"ya istinaden) "oks-fos savaşları" olarak bilinir. Çekişmeler, ancak Mitchell'ın 1978'de Nobel Ödülü almasıyla sona ermiştir.
Burada bir ironi var. Mitchell'ın Nobel'i kimya dalındaydı; ama düşünceleri aslında kimyanın elenmesine ilişkindi. Nasıl ki genetik kod, enformasyonun kimyayı aşmasını sağladıysa, Mitchell'ın proton değişim dereceleri de, hücresel metabolizmanın kimyayı aşmasını sağlıyordu. Proton değişim derecelerinin (proton gradyanlarının) kullanılması, yaşamın başlangıcına ilişkin bir anlayış sunmakla kalmayıp, 4 milyar yıllık evrimsel süreçte sadece bir kez gerçekleşen karmaşık ökaryotik (çekirdekli) hücrelerin evrimi olayına da ışık tutuyordu.
Hücrelerin Nefes Alışı
Hücrelerimiz besini oksijen ile yakar ve ATP (yaşamın evrensel enerji birimi) biçiminde salınan enerjiyi korumayı başarır; bu sürece aerobik solunum denir. Hücreler bunu nasıl yapar ve nasıl yapmayabilirler?
1940'lı yıllarda Efraim Racker hücrelerin oksijen yokluğunda, glukozun parçalanmasından az miktarda enerji topladığı mekanizmanın farkına vardı. Glikoliz adı verilen bu olayda, ATP oluşturmak için şeker moleküllerinden fosfat grupları doğrudan ADP'ye aktarılır. Tüm süreç saf kimyadır. Bir molekülün diğeriyle tepkimesi ve böylece stokiyometri yasalarına uyması söz konusudur. Yani denklemleri dengeleyebilirsiniz. Bekleneceği üzere, Racker ve arkadaşları derhal bu kavrayışlarını, niceliksel olarak çok daha önemli bir süreç olan aerobik solunuma aktarmayı denemişlerdir. Aerobik solunum, ATP'mizin %80'den fazlasını sağlar.
Fakat aerobik solunumla ilgili bariz sorunlardan biri, dengelenmemesidir. Tüketilen her oksijen molekülü başına tam olarak ne kadar ATP üretilir? Miktar değişir ama yaklaşık olarak 2,5 ATP molekülü civarındadır. Glukoz başına ATP miktarı ise yine değişken olmakla beraber 28-38 arasındadır. Aerobik solunum stokiyometrik değildir; dolayısıyla aslında kimya değildir. İşte yüksek enerjili bir kimyasal aracı (glukozun oksidasyoundan gelen enerjiyi ATP oluşturmak için aktarabilen bir molekül) bulma çabalarının başarısızlığa mahkum olmasının nedeni budur.
Böyle bir aracı yerine, Mitchell zardan geçen bir proton değişim derecesi (proton gradyanı) önermiştir: Proton harekete geçirme kuvveti. Hidroelektrik bir baraja benzer şekilde çalışır. Besin oksidasyonu ile salınan enerji, protonları zardan (baraj) karşıya pompalamak için kullanılır. Böylece etkisel olarak, zarın bir tarafında bir proton deposu oluşur. Bu zara gömülü olan şaşırtıcı protein türbinlerinden proton akışı, ATP sentezine güç sağlar; tıpkı mekanize türbinlerden akan suyun elektrik üretmesi gibi. Bu, solunumun neden stokiyometrik olmadığını açıklar: Bir derecelilik (gradyan), yapısı gereği derecelenmelerden oluşur.
İnce Ayrıntılar
Mitchell bazı ayrıntılar konusunda bütünüyle yanılıyordu ama genel kavrayışı doğruydu; hatta devrimseldi. Sözcüğün gerçek anlamıyla devrimseldi, çünkü ATP sintaz enzimi devir yapıyordu. Protonların zar türbinlerinden akışı, ATP sintazın sapını döndürüyor ve bu dönüşle tetiklenen uyuşumsal değişimler, ATP sentezini katalizliyor. Bu mekanizma ilk olarak Paul Boyer tarafından önerilmişti. Kendisi uzun süre boyunca mekanikler konusunda Mitchell'ın fikirlerini kabul etmedi ve bunda haklı olduğu sonradan John Walker tarafından x-ışın kristalografisiyle kanıtlandı. Boyer ve Walker, 1997 yılında Nobel Ödülü'nü paylaştı.
Son 20 yılda gerçekleşen moleküler biyoloji alanındaki başarılar, 2010 yılında doruğa çıktı. Bir başka solunumsal kompleks olan devasa (bir proteine göre) kompleks I'in kristal yapısı, Leonid Sazanov tarafından çözüldü. Bir kez daha yapı, mekanizmayı ele verdi; bu kez dönel bir motor değildi ama daha da şaşırtıcı şekilde, bir buhar makinesinin pistonundan hiç de farklı olmayan bir kaldıraç mekanizmasıydı.

Kompleks I'in x-ışın kristalografisi ile ortaya çıkarılan yapısı.
Yapı (a), gösterilen piston mekanizmasına (b) işaret ediyor.
Pistonun yerini değiştirmek, protonları üç ayrı kanaldan zarın karşısına itiyor.
© 2010 Nature Publishing Group Efremov, R. G., Baradaran, R., & Sazanov
Bu ustaca başarıların yerilecek bir tarafı olmamakla birlikte, Mitchell'ı ilk olarak harekete geçiren sorular, şaşırtıcı biçimde yanıtlanmadan kaldı. Solunumun nasıl işlediğini neredeyse atomik ayrıntısıyla biliyoruz. Neden o şekilde işlediği hakkındaki bilgimiz ise çok daha az.
Proton Harekete Geçirme
Mitchell mitokondriler üzerinde çalıştı; çünkü bunu yapabilirdi. Mitokondriler, kolay izlenebilir bir deneysel modeldi. Ama soruya bakteriyel fizyolojinin bakış açısından yaklaştı: Bakteriler içlerini nasıl dışarıdan farklı tutar? Mitchell yaşamı boyunca solunumun ayrıntılı mekanizmasını bu çok daha geniş olan açıdan gördü: Zar proteinleri, zardan öteye gradyanlar yaratabilir ve bu gradyanlar da işe güç sağlayabilir. ATP sentezine güç sağlayan proton gradyanları, Mitchell için sadece bir özel durumdu.
Protonların baskın rolünü pek tahmin edememişti. Hücreler her ne kadar sodyum, potasyum ya da kalsiyum gradyanları da oluştursalar, proton gradyanları daha önemliydi. Protonlar, mitokondrilerin yanı sıra bakteriler ve arkelerde de solunuma güç sağlıyordu. Proton gradyanları, tüm fotosentez biçimlerinde merkezi rol oynuyordu; ayrıca bakteriyel hareketlilik (ATP sintaza benzeyen dönel bir motor olan ünlü flagellar motor yoluyla) ve homeostasisde de (çoğu molekülün hücreye giriş-çıkışı doğrudan proton gradyanı ile bağlantılıdır) öyle. ATP üretmek için proton gradyanlarına gereksinimleri olmayan mayalandırıcılar bile proton pompalamaya güç sağlamak için mayalanmadan elde edilen ATP'yi kullanarak, proton harekete geçirme kuvvetini sürdürür.
Kısacası, Mitchell protonların öneminin farkındaydı ama bu önemin derecesini pek tahmin edemedi. Peki ama neden protonlar? Çünkü onlar en baştan beri oradalardı, diyor NASA'dan yerkimyacı Michael Russell.
Canlılığın Başlangıcındaki Proton Gradyanları
Yaklaşık son 20 yıldır, yaşamın kökenine ilişkin anlayışımızda beliren yaklaşım değişiminde (paradigma kaymasında), Russel etkin bir kuvvet oldu. Cevher jeokimyası (çoğu cevher, hidrotermal baca sistemleri tarafından çöktürülür) geçmişi olan Russell, orta-Atlantik'teki modern Kayıp Şehir baca sistemine benzeyen alkalin bacaların, yaşam için ideal kuluçkalar olabileceğini öne sürdü. Bunlar sürekli olarak hidrojen gazı, karbon dioksit, mineral katalizörler ile birbirlerine bağlı mikro-gözenek labirenti (film benzeri zarlarıyla, hücrelere benzeyen doğal bölümler) sağlayabilirdi.

Bunlar volkanik olmayan serpantinleşme süreci ile oluşmuştur.
© 2001 Nature Publishing Group Kelley D. S. et al.
Alkalin bacalar, özünde, denge durumundan çok uzak bir durumda işleyen elektrokimyasal reaktörlerdir. Ama Russell'ın düşüncesinin en önemli öğesi, doğal proton gradyanlarında yatar. 4 milyar yıl önce alkalin akışkanlar, hafif asidik olan (CO2 düzeyleri bugünkünden yaklaşık bin kat daha yüksekti ve CO2 çözeltide karbonik asit oluşturarak, okyanusları hafif asidik bir hâle getiriyordu) okyanuslarda kabarcıklar oluşturuyordu. Asidiklik, proton derişiminin bir ölçüsüdür ve bu derişim okyanuslarda, baca akışkanlarında olduğundan yaklaşık dört mertebe (dört pH birimi) daha yüksektir. Aradaki bu fark, modern hücrelerin zarları ile aynı polariteye (dışı pozitif) ve benzer elektrokimyasal potansiyele (yaklaşık 200 mV) sahip olan baca zarları boyunca doğal bir proton gradyanına neden olur.
Russell uzun süredir, doğal proton gradyanlarının, canlılığın başlamasına güç sağlamada merkezi bir rol oynadığını iddia ediyor. Elbette yanıt bekleyen büyük sorular var. Örneğin, ATP sintaz gibi yüksek teknolojili bir protein mekanizmasından yoksun olan ilk hücreler, gradyanları nasıl yüklemiş olabilir? Şu anda incelenen birkaç olası abiyotik mekanizma bulunuyor. Termodinamik görüşler ise canlılığın başlayabilmesi için tek yolun, proton gradyanlarını yüklemenin bir yolunun bulunması olabileceğini belirtiyor.
Proton Gradyanları Neden Gerekli?

Substrat düzeyinde fosforilasyon,
1 ATP kazanmak için 1 ATP harcar.
Net büyüme olanaksızdır.
© 2010 Nature Education All rights reserved.
Proton gradyanlarının gerekme nedeni kimyaya indirgenebilir. Yaşam, karbon dioksiti hidrojenler. Diğer bir deyişle, karbon dioksiti organik moleküllere dönüştürmek için canlılık CO2'ye hidrojen atomları tutturur. Bunu yapmanın çok fazla yolu olmasına karşın, tüm canlılık sadece beş ana yol kullanır. Bunlardan biri dışında hepsi enerji harcar (örneğin, fotosentezde güneş enerjisi kullanılır). İstisnai durum ise antik bir yol olan "asetil-CoA" sürecidir. Bu süreçte hidrojen gazı bir dizi basamak yoluyla karbon dioksit ile tepkimeye girer. Bu süreç, hücre metabolizmasındaki en önemli moleküllerden biri olan pirüvata kadar egzotermiktir (yani ATP olarak yakalanabilecek enerji salar). Everett Shock'un sözleriyle bu süreç, yemeniz için üzerine para verilen ücretsiz bir yemek gibidir.
Ama Russell ile birlikte çalışan William Martin tarafından dikkat çekilen bir sorun var. Günümüzde asetil-CoA yolunu kullanan tüm hücreler, proton gradyanlarına bağlıdır. Hiçbiri mayalanma (yani glikoliz kimyası) ile büyüyemez. Peki neden? Çünkü CO2 kararlı bir moleküldür ve hidrojene bile kolay kolay tepki vermez; termodinamik tepki vermesini söylese bile. Karbon dioksit bu açıdan biraz oksijene benzer: Tepki vermeye bir başladı mı, kolayca durdurulmaz; ama başlaması için bir kıvılcıma gerek vardır. Hücrelerin karbon dioksitin tepki vermeye başlaması için gerek duyduğu kıvılcım ATP'dir. Sorun şu ki, CO2 ile H2 tepkimesi enerji salar ama çok değil; sadece tek 1 ATP yapmaya yetecek kadar. Bunun anlamı, hücrelerin 1 ATP kazanmak için 1 ATP kaybetmeleri gerektiğidir. O halde net bir kazanç yoktur. Kazanç yoksa, büyüme de yoktur ve büyüme olmazsa yaşam da olmaz.

Gradyanlar bu döngüyü kırar. CO2 ile H2 tepkimesinin 1 ATP yapmaya yetecek enerji saldığı tam olarak doğru değildir; aslında 1,5 ATP yapmaya yetecek kadar enerji salınır. Ama elbette 1,5 ATP diye bir şey yoktur; en azından stokiyometrik kimyada. Dolayısıyla tepkimeden artan enerji kimyasal açıdan yitirilir. Ama gradyanda böyle olmaz. İlkesel olarak, bir protonu zardan dışarı pompalamak için bir tepkime tekrar tekrar yinelenebilir. Yeterince proton biriktiğinde, proton hareket etme kuvveti ATP oluşumunu başlatır. Böylece bir gradyan, hücrelerin "bozuk para" olarak proton biriktirmesine olanak tanır. İşte dünyadaki tüm farkı yaratan budur.
Karmaşık Yaşamın Kökeni
Güçlerine rağmen protonlar da sorunlardan kendilerine düşen payı alır. Bu sorunlar, canlılığın neden 2 milyar yıl boyunca bir tekdüzeliğe sıkışıp kaldığını açıklayabilir. Günümüzde Dünya üzerinde bulunan tüm karmaşık yaşam türleri, belli bir karmaşık hücre tipinden oluşur: Ökaryotik hücre. Genellikle bakterilerden ve arkelerden çok daha iri olan ökaryotik hücrelerde bir çekirdek bulunur. Genomları da daha büyük olur ve ayrıca çok çeşitli özelleşmiş organellere (minik organcıklara) sahiptirler.
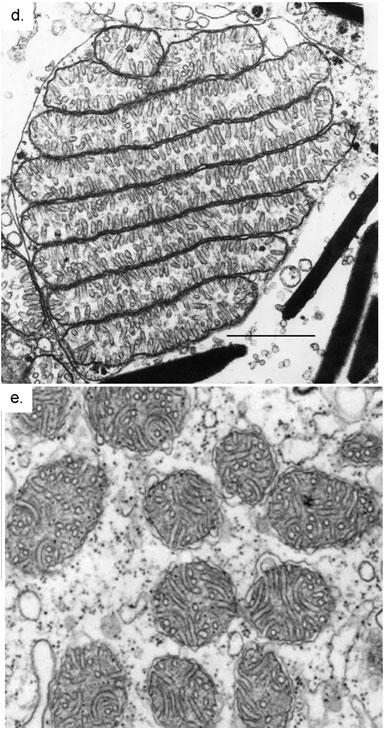
© 2010 Nature Education Courtesy of
Mark Farmer (d) & Richard Allen (e).
İlginç olan şu ki, ökaryotlar sürekli olarak bitki, hayvan, mantar ve alg gibi büyük, karmaşık, çok hücreli organizmalar oluşturma eğiliminde olmuştur. Prokaryotlar ise daha büyük morfolojik karmaşıklığa evrilme yönünde çok az eğilim gösterir; biyokimyasal ustalıklarına rağmen. Peki neden? Olası yanıtlardan biri proton gradyanlarının kontrolü ile ilişkilidir. Tüm ökaryotik hücrelerin mitokondrilerinin olduğu ya da en azından bir zamanlar sahip olup, sonradan yitirdikleri anlaşılmıştır. Mitokondri olmadan ökaryot olmaz. Oksidatif fosforlama becerisi olan tüm mitokondrilerde, kendilerine ait ufak bir genom bulunur. Görünüşe bakılırsa bu genom, zar potansiyeli üzerinde kontrol sağlayabilmek için gereklidir. 5 nanometrelik zar boyunca 150 mV'luk bir zar potansiyeli, metre başına 30 milyon voltluk bir alan gücü verir; bu bir yıldırımınkine eşdeğerdir.
Bu devasa elektrokimyasal potansiyel, mitokondriyal zarları, hücre içindeki tüm diğer zar sistemlerinden bütünüyle farklı kılar. Allen'a göre, hücresel alt-bölgelerde yerel olarak mitokondriyal genlerin bulunması, bu nedenle gereklidir. Etkisel olarak, elektrokimyasal potansiyeldeki yerel değişikliklere yanıt vererek, hücrenin kendi kendini elektrikle çarpmasını önler. Mitokondriyal genom olmazsa, oksidatif fosforlama da olmaz. O halde bakterilerin hücre ve genom büyüklüklerini genişletememelerinin nedeni, enerjili zarları ile doğru gen kümelerini fiziksel olarak eşleştirememeleri olabilir. Mitokondriler olmadan bakteriler büyüyemez ve karmaşıklaşamaz, çünkü geniş bir enerjili zar alanı boyunca solunumu kontrol edemezler. Eğer durum buysa, mitokondrilerin edinimi ile karmaşıklığın kökeninin aslında aynı olay olduğu söylenebilir.
- Scitable, "Why Are Cells Powered by Proton Gradients?" https://www.nature.com/scitable/topicpage/why-are-cells-powered-by-proton-gradients-14373960
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek OlBunlar da İlginizi Çekebilir



En Çok Okunan
Bu Ay Öne Çıkanlar



İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.