

Öz
Arkaplan
450 milyon yıldan daha uzun süre önce, bir deniz kordalısı (sırt ipliklisi) soyunda, omurgalıların ortak atasına varmadan önce, iki "tüm genom ikilenmesi"nin (TGİ ) ardı ardına gerçekleştiği ileri sürülmüştür. Temel teşkil eden bu olayların eksiksiz bir şekilde yeniden yapılandırılması, bu eski TGİ'lerin, varolan omurgalılar üzerindeki etkisini daha iyi anlamak için bir çerçeve sağlar.
Bulgular
Omurgalı evriminin başlangıcındaki kromozomların evrimini yeniden yapılandırdık. 326 milyon yıllık atasal bir Amniota genomunda bulunan genlerin oldukça bitişik durumdaki sıralanışını yeniden yapılandırmak için varolan 61 hayvanın genomlarını karşılaştırarak çalışmaya başladık. Bu genomda, ikilenmiş kromozomların tetradlarını belirlemek için iki TGİ'den kaynaklanan ikilenmiş genlerin iyi-desteklenmiş bir listesini oluşturduk. Bunu kullanarak, 17 kromozomlu bir omurgalı-öncesi genomunun ikilenerek 34 kromozoma çıktığı ve yedi kez kromozom kaynaşması geçirdikten sonra yeniden ikilenerek, 54 kromozomlu olduğu bir kronoloji yapılandırdık. Gnathostomata (çeneli omurgalılar) soyunun Cyclostomata (varolan çenesiz balıklar) soyundan ayrılmasından sonra, dört kaynaşma daha gerçekleşerek, 50 kromozomlu atasal Euteleostomi (kemikli omurgalılar) genomunu oluşturdu.
Çıkarımlar
Bu bulgular, omurgalıların atasından önce varolmuş soyda iki TGİ'nin gerçekleştiğini sağlam bir şekilde ortaya koyarak, bufa (taş emen) balığının genomunun çözümlenmesiyle ortaya çıkan belirsizliği de çözüme kavuşturuyor. Bu çalışma, bir ortak ata yönünden omurgalı kromozomlarının evriminin ve özellikle de varolan omurgalı genomlarının gen bileşimlerinde ortaya çıkan ikilenmiş gen muhafazası ve kaybının örüntüsünün incelenmesi için bir temel sağlıyor.
Arka Plan
Yeni gen kopyaları, genom evrimi sırasında küçük ölçekli ikilenmelerle bolca belirerek, genetik yeniliklere ve fenotipik çeşitliliğe katkı yapar. Omurgalı evriminde, tüm genom ikilenmesi (TGİ) enderdir; bitkilerde ise oldukça sık gibi görünmektedir. 450 milyon yıldan uzun süre önce, eski bir omurgalı soyu, varolan yaklaşık 60.000 türe çeşitlenmesinden önce nispeten kısa aralıklı olarak iki tane TGİ geçirdi. 1970'li yılların başından bu yana, Susumu Ohno'nun gözünde canlandırdığı ve "1R-2R hipotezi" olarak bilinen bu olaylar, genom çapında yapılan çeşitli çalışmalarla sağlam bir şekilde ortaya kondu.
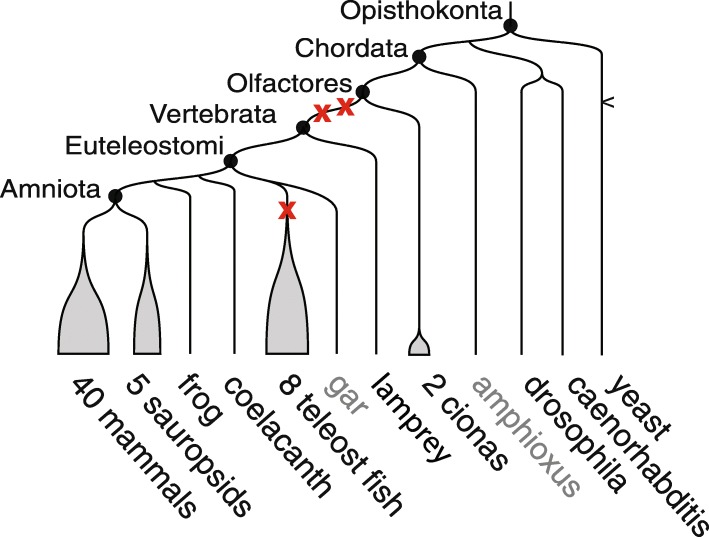 FIG.1: Bu çalışmada kullanılan türler arası ilişkileri gösteren filogenetik şema. Dalların ucunda siyah yazılan türler veya tür grupları, Ensemble sürüm 69'a dahil edilen 61 türdür ve atasal Amniota genomunun yeniden yapılandırımı için kullanılmıştır. Sauropsid'lere kuşlar ve sürüngenler dahildir. Gri yazılan türler, çözümlemenin başka aşamalarında kullanılmıştır. Siyah daireler, bu çalışmayla ilgisi bulunan atasal genomları belirtir. Kırmızı çarpılar, TGİ'lerin göreli konumlarına işaret eder: omurgalı yayılımında önce iki tane ve teleost balık yayılımında önce bir tane. Dallar ölçeksizdir. FIG.1: Bu çalışmada kullanılan türler arası ilişkileri gösteren filogenetik şema. Dalların ucunda siyah yazılan türler veya tür grupları, Ensemble sürüm 69'a dahil edilen 61 türdür ve atasal Amniota genomunun yeniden yapılandırımı için kullanılmıştır. Sauropsid'lere kuşlar ve sürüngenler dahildir. Gri yazılan türler, çözümlemenin başka aşamalarında kullanılmıştır. Siyah daireler, bu çalışmayla ilgisi bulunan atasal genomları belirtir. Kırmızı çarpılar, TGİ'lerin göreli konumlarına işaret eder: omurgalı yayılımında önce iki tane ve teleost balık yayılımında önce bir tane. Dallar ölçeksizdir.
FIG.1: Bu çalışmada kullanılan türler arası ilişkileri gösteren filogenetik şema. Dalların ucunda siyah yazılan türler veya tür grupları, Ensemble sürüm 69'a dahil edilen 61 türdür ve atasal Amniota genomunun yeniden yapılandırımı için kullanılmıştır. Sauropsid'lere kuşlar ve sürüngenler dahildir. Gri yazılan türler, çözümlemenin başka aşamalarında kullanılmıştır. Siyah daireler, bu çalışmayla ilgisi bulunan atasal genomları belirtir. Kırmızı çarpılar, TGİ'lerin göreli konumlarına işaret eder: omurgalı yayılımında önce iki tane ve teleost balık yayılımında önce bir tane. Dallar ölçeksizdir. FIG.1: Bu çalışmada kullanılan türler arası ilişkileri gösteren filogenetik şema. Dalların ucunda siyah yazılan türler veya tür grupları, Ensemble sürüm 69'a dahil edilen 61 türdür ve atasal Amniota genomunun yeniden yapılandırımı için kullanılmıştır. Sauropsid'lere kuşlar ve sürüngenler dahildir. Gri yazılan türler, çözümlemenin başka aşamalarında kullanılmıştır. Siyah daireler, bu çalışmayla ilgisi bulunan atasal genomları belirtir. Kırmızı çarpılar, TGİ'lerin göreli konumlarına işaret eder: omurgalı yayılımında önce iki tane ve teleost balık yayılımında önce bir tane. Dallar ölçeksizdir.1R-2R TGİ'lerinden dolayı, varolan insan genlerinin yaklaşık %35'i hâlâ ikilenmiş kopyalar (ohnologlar) hâlindedir; buna bütün tetrapodlarda bulunan dört küme HOX geni de dahildir. Ohnologlar, insan genomundaki ikilenmiş genlerin büyük çoğunluğunu temsil eder. Hastalık genlerinde bol bulundukları ve insan topluluklarındaki yapısal vasryasyonların sıklığını etkiledikleri gösterilmiştir. O hâlde antik 1R-2R genom ikilenmeleri, hâlâ bugünkü genomlar üzerinde güçlü bir etki yaparak, onların eski tarihlerinin daha iyi anlaşılmasını icap ettirir. 1R-2R'den önce, 1R-2R sırasında ve 1R-2R'den sonra karyotiplerin evriminin ilk yeniden yapılandırılması, çok sayıda soruyu yanıtsız bırakmıştır. Tüm atasal omurgalı kromozomlarını ve iki TGİ arasında kromozom kaynaşmalarının ya da ayrışmalarının olup olmadığını yanılma payı olmaksızın belirleyememiştir. Bu düşük çözünürlüğün nedeni, omurgalı atadan (Vertebrata) bu yana kromozomların çok kez yeniden düzenlenerek, varolan genomlardaki atasal gen organizasyonunu karıştırması ve uygun bir dış-grup türün yokluğudur. Buna ek olarak, bufa balığı genomu ile tavuk genomu arasında yakın zamanda yapılan bir karşılaştırma, 1R-2R hipotezini sorgulamış ve tek bir TGİ ile çok sayıda kısmi ikilenmenin de bu iki tür arasında gözlemlenen aynı gen sıralı örüntüleri açıklayabileceğini ortaya atmıştır. Hem 1R-öncesi genomun ilk yeniden yapılandırmasındaki kesinsizlikler, hem de 1R-2R hipotezinin bu alternatifi, omurgalı kromozomlarının erken dönem evrimlerinin ayrıntılı bir çözümlemesini gerektirmiştir.
Dizilenmiş omurgalı genomlarının sayısı son zamanlarda çok artarak, atasal genomların daha yüksek bir doğrulukla yeniden yapılandırılmasına olanak tanıdı. Erken dönem omurgalı karyotip evriminin ayrıntılarını daha iyi anlamak için bir atlama taşı olarak atasal Amniota genomunu yeniden yapılandırdıktan sonra, 1R-öncesi omurgalı genomunun 17 kromozom içerdiğini ve tüm omurgalıların 54 kromozom içeren bir 2R-sonrası genomdan türediğini bulduk. İnsan genomunun hâlâ 1R-öncesi genomun güçlü bir izini taşıdığını gösterdik ve tüm ardıl türler tarafından paylaşılan bir referans noktasının perspektifinden, omurgalılarda iki TGİ'nin etkisini incelemek için kaynaklar sağladık.
Bulgular
Atasal Amniota genomundaki ohnolog gen çiftlerinin belirlenmesi
Kuşların, sürüngenlerin ve memelilerin (326 milyon yıl) atası olan atasal Amniota genomunda 19.786 gen bulunduğunu, Ensembl gen ağaçlarından (versiyon 69) çıkarsadık. Bu genleri Amniota genomunda bulundukları şekilde sıralayıp yönlendirmek amacıyla AGORA (Algorithm for Gene Order Reconstruction in Ancestors) kullandık. Bu in silico (bilgisayar ortamında) gerçekleştirilen yeniden yapılandırma 470 bölümden oluşuyordu; bölümlerdeki genlerin %50'si 253 genden (N50 uzunluğu) daha büyüktü. Daha sonra ikilenmiş bölgeleri belirlemek için bir Bitişik Atasal Bölgeler (BAB ; ortalama BAB uzunluğu 256 gen, toplam 12.134 gen) başlangıç kümesi olarak 50 genden daha büyük olan 56 tane kromozom-boyutlu bölüm seçtik.
İkilenmiş kromozom bölümleri çiftlerinin belirlenmesinde, 1R-2R TGİ'lerinden dolayı oluşan ohnologlar kilit rol oynar. İkilenmelerini tarihlemek için gen ağaçları kullanarak, varsayımsal ohnolog genleri doğrudan yeniden yapılandırılmış Amniota genomunda belirledik; bunu yaparken bir çiftin her bir üyesinin farklı bir BAB'a ait olduğundan emindik. Böylece 5616 atasal Amniota ohnolog çifti içeren "Liste A"yı oluşturduk. Ayrıca daha önce yapılan iki çalışmada, insanda ve başka omurgalı genomlarında, korunmuş aynı gen sıralaması ve dizilim benzerliği kullanılarak, iki TGİ'nden dolayı oluşan ohnologlar belirlenmişti. Bu çalışmalardan çıkan varolan gen belirleyicilerini, onların atasal Amniota gen belirleyicilerine dönüştürmek için biz Ensembl gen ağaçlarını kullandık.
Makino ve McLysaght tarafından ortaya konan ve buradan itibaren "Liste B" olarak adlandırılacak olan ilk liste, 4870 atasal Amniota gen çifti içermektedir. Singh ve ark. tarafından yapılan ikinci çalışma, ohnologları tanımlamak için üç güvenilirlik düzeyi (sıkı, orta ve gevşek) ortaya koymuştur. Bu ölçütlere uyarak, biz üç ek liste daha tanımladık; bunlar sırasıyla 2873 (Liste “C-sıkı”), 5253 (Liste “C-orta”) ve 7806 (Liste “C-gevşek”) atasal Amniota ohnolog çifti içermektedir.
A, B ve C-gevşek listelerinin toplamı, genlerin sadece %25'inin ortak olduğunu göstermektedir ama biz üç listenin buna rağmen 1R-2R hipotezini desteklediğini aşağıda gösterdik. Bu senaryoda, her bir orijinal kromozom önce iki sonra dört kopyaya ikilenmiştir; böylece kromozomlar tetradlar oluşturur ve her biri üç ohnolog tamamlayıcıya sahip olur. Beş adet ohnolog listesinin her birini kullanarak, BAB çiftlerinin, eğer rastgele dağıtılsalar, beklenenden daha fazla ohnolog paylaşıp paylaşmayacaklarını sınadık (orantısallık testi). Bütün durumlarda, BAB'ların ortalama olarak diğer üç BAB ile ohnolog olduklarını gösterdik. O hâlde, farklılıklarına karşın tüm listeler 1R-2R hipotezini destekleyerek, beş listenin hepsi kullanılarak yapılandırılacak gelişmiş bir uzlaşımsal atasal Amniota ohnolog çiftleri listesini gerekçelendirir. En güvenilir altküme (1273 ohnolog çifti) olarak A, B ve C-sıkı listelerinin kesişiminde başladık ve düşük güvenilirlikli altkümelerden (C-orta ve C-gevşek içeren listeler veya daha az listenin kesişimindeki gen çiftleri) gen çiftleri ekleyerek aşamalı olarak bunu genişlettik. Bu süreçte, büyümekte olan listenin 1R-2R hipotezi ile uyumlu kaldığından emindik: eğer iki gen farklı filogenetik ağaçlara aitse çift asla dahil edilemezdi (yani bir çiftteki ikilenmiş iki gen, ortak bir atasal genden türemiş olmalıdır) ve bir filogenetik ağaç içindeki bütün ohnologlar, çiftler şeklinde düzenlendiğinde, dörtten (bir tetrad) fazla BAB'ı bağlantılandıramaz. Bu kademeli süreç, 8184 ohnolog genden oluşan bir liste verdi; 7441 ohnolog çift, 2973 ohnolog ailesi içinde gruplandırıldı; her bir aile, ilkesel olarak, bir tane 1R-öncesi gene karşılık geliyordu.
 FIG.2: Atasal Amniota genomundaki ohnolog çiftlerin belirlenmesi. (a) Beş adet Amniota ohnolog çifti listesi arasındaki karşılaştırma. (b) Ohnolog çift seçiminin şematik örneği. FIG.2: Atasal Amniota genomundaki ohnolog çiftlerin belirlenmesi. (a) Beş adet Amniota ohnolog çifti listesi arasındaki karşılaştırma. (b) Ohnolog çift seçiminin şematik örneği.
FIG.2: Atasal Amniota genomundaki ohnolog çiftlerin belirlenmesi. (a) Beş adet Amniota ohnolog çifti listesi arasındaki karşılaştırma. (b) Ohnolog çift seçiminin şematik örneği. FIG.2: Atasal Amniota genomundaki ohnolog çiftlerin belirlenmesi. (a) Beş adet Amniota ohnolog çifti listesi arasındaki karşılaştırma. (b) Ohnolog çift seçiminin şematik örneği.Birkaç nedenden ötürü, ohnolog çiftler listesi yüksek kalitededir. Birincisi, yeniden yapılandırılmış atasal Amniota genomundaki aynı gen sıralamasına ve gen içeriğine dayanır; bu genom, 1R-2R olaylarına, varolan genomlardan 326 milyon yıl daha yakındır; dolayısıyla 1R-2R olaylarının izleri daha büyük bir doğrulukla okunur. İkincisi, bu liste bir 1R-2R uyumluluk kuralına bağlı kalmıştır; yani hiçbir ohnolog gen ailesi dörtten fazla BAB bağlantılandırmaz. Üçüncüsü, ohnolog çiftlerin hepsi filogenetik olarak tutarlıdır; bir çiftteki her iki gen daima aynı Ensembl gen ağacına aittir. Dördüncüsü, genlerin birlikte ikilendiğine ilişkin yanlış düşüncelerden kaçınmak için bir çiftin iki geninin, ancak ≥ 90 gen onları ayırıyorsa aynı BAB üzerinde olmalarına izin verilmiştir.
2R-sonrası ikilenmiş BAB'ların belirlenmesi
Geliştirilmiş ohnolog çiftler listemizi kullanarak, 2R-sonrası karyotipe mümkün olduğunca benzer bir konfigürasyona dönüştürmek amacıyla atasal Amniota BAB'larını manuel olarak ayırdık, düzenledik ve grupladık. En basit senaryoda, 2R-sonrası BAB'lar hemen, her biri bir 1R-öncesi kromozoma karşılık gelen dört ohnolog BAB'lı tetradlar oluşturmuş olmalıdır. Ancak 1R ile 2R arasında veya 2R ile Amniota arasında kromozomların yeniden düzenlenmeleri ve BAB'ların eksik ya da hatalı yeniden yapılandırımları hep birlikte elbirliğiyle ideal örüntüyü bozar. En büyük 56 BAB ile başladık ve yukarıda tanımlandığı şekilde dikkate değer sayıda ohnolog gen paylaşan BAB'ları (yani ohnolog BAB'ları) belirlemek için orantısallık testi uyguladık. Hepsi de dikkate değer ölçüde ohnolog çiftlik gösteren en az üç BAB grubu belirledik (p değeri < 5.10−2, Bonferroni ayarlı). Bunlar, daha küçük BAB'lar ve/veya düşük dikkate değerlik eşiğindeki BAB'lar dahil edilerek, tetrad şeklinde tamamlandı (yani dört BAB'ın hepsi dikkate değer ölçüde birbiriyle ohnolog). Ayrıca, aynı Amniota kromozomuna ait olduğuna ilişkin kanıt görünen BAB'ları da birleştirdik, çünkü alternatif AGORA yeniden yapılandırımlarında bunlar farklı parametre kümeleri kullanılarak birleştirilmişlerdi ve/veya Amniota ardılı genomlarında (insan veya tavuk) bunlar özdeş homolojiler sergiliyordu. Ek olarak, birleştirilmiş BAB'lar, bir üçlüdeki en azından bir ortak BAB'la dikkate değer ölçüde ohnolog olmalıydı ve birbirleriyle dikkate değer bir ohnologluk göstermemelilerdi. Ayrıca, uzunlukları boyunca ohnologların dağılımında bozukluk gösteren ve onların tavuk, insan, benekli zargana veya Japon pirinç balığı kromozomlarına homolojilerinde bozukluk gösteren BAB'ları ayırdık. Son olarak, benekli zargana veya Japon pirinç balığı gibi dış-grup türlerle homolojileri kullanarak, birleşmiş BAB'ları doğruladık.
 TABLO: Amniota BAB'larının düzenlenmesi. TABLO: Amniota BAB'larının düzenlenmesi.
TABLO: Amniota BAB'larının düzenlenmesi. TABLO: Amniota BAB'larının düzenlenmesi.Amniota BAB'larının, özellikle de insan kromozomlarına kıyasla, bir 2R-sonrası konfigürasyona bu dönüşümünden sonra BAB'ların tetradlara ayrılması daha net hâle geldi. İnsan kromozomları p değeri eşiğine çok daha fazla duyarlı iken, orantısallık testi herhangi bir p değeri eşiğinden neredeyse bağımsız olarak, her bir düzenlenen Amniota BAB'ını ortalama 3 başka BAB ile bağlantılandırdı. Aslında insan genomunda, kromozom başına üç partnerin beklenen ortalamasına sadece 1.10−09 şeklinde bir p değerinde ulaşılır; 7 insan kromozomunun bir tetrada ayrılamayacağı bir yerde zorlayıcı bir eşik. Bir tetradın yapılandırılmasına ve BAB'ların düzenlenmesine bir örnek Fig. 3c'de ayrıntılandırılmıştır. Bir üçlüde gruplanan dikkate değer ölçüde ohnolog üç BAB belirledik (73, 117, 250 BAB'ları). İki başka BAB da (256 and 137) BAB 250 ile dikkate değer bir ohnologluk sergiliyor. Dikkate değer ama daha yüksek p değeriyle, başlangıçtaki beş BAB'ın dördüyle bağlantılı olan daha küçük bir BAB'ın (BAB 82) eklenmesiyle tetrad tamamlandı. Ardından, başlangıçtaki beş BAB'ın üçü, daha büyük tek bir BAB'da düzenlenmek için koşulları sağladı (117, 137, 256 BAB'ları): Bunlar ortak BAB'larla ohnolog idi ama birbirleriyle ohnolog değildi ve daha yeni bir Ensemble versiyonu (Versiyon 84) kullanınca, AGORA 117/137 BAB'ları ile 137/256 BAB'ları birleştirdi. Bundan başka, üç BAB'ın aynı tavuk ve benekli zargana kromozomlarına haritalama yapması, ortak Vertebrata atalarının aynı kromozomundan türediklerine güçlü biçimde işaret etti. Son olarak, düzenlenen BAB'ların üçü de Japon pirinç balığı (ilave bir TGİ daha geçirmiş olan bir teleost balık) genomu üzerine haritalandığında, aynı iki Japon pirinç balığı kromozomu (13 ve 14) ile ortologdu.
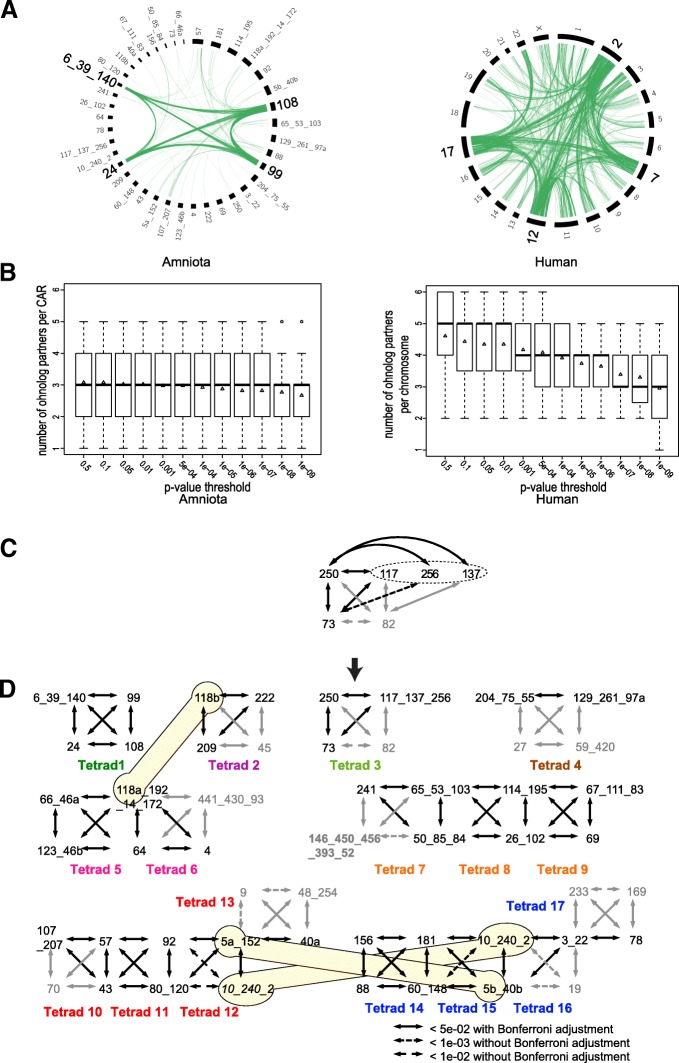 FIG.3: Atasal Amniota BAB'larıın tetradlarda organizasyonu. FIG.3: Atasal Amniota BAB'larıın tetradlarda organizasyonu.
FIG.3: Atasal Amniota BAB'larıın tetradlarda organizasyonu. FIG.3: Atasal Amniota BAB'larıın tetradlarda organizasyonu.İki Amniota BAB'ının her birinin iki alt-BAB'a (5 ve 118 BAB'ları) ayrılmasını gerektiren iki 2R-sonrası kromozomal kaynaşma belirledik ve üç BAB'ın (40, 46 ve 97 BAB'ları) ayrılmasını gerektiren üç olası düzenleme hatası belirledik. Ayrıca, 2R-sonrası konfigürasyonlarını daha yakın temsil edecek şekilde düzenlenen 51 BAB'lık bir son küme ile biten 23 BAB düzenlemesi gerçekleştirdik. Orantısallık testinin dikkate değer p değerlerine dayalı olan BAB'lar arası ohnoloji ilişkileri, 51 BAB'ı 17 tetrad şeklinde bağlantıladı. BAB'ları ayırmak veya düzenlemek ve onları tetrad şeklinde gruplamak için izlediğimiz eksiksiz adım adım prosedür, ek dosyalarda ayrıntılandırıldı.
1R ve 2R tüm genom ikilenmeleri arasındaki kromozom evrimi
51 Amniota BAB'ından oluşan 17 tetradın hepsi ayrık değildir: bazıları ortak bir veya iki BAB paylaşarak, 1R ile 2R arasındaki ve 2R'den sonraki kromozomal olayları yansıtır. İlk olarak her bir konfigürasyona karşılık gelen kuramsal senaryoları ortaya koyarak, bu olayları belirledik. Ayrık olmayan tek bir tetrad, iki TGİ arasında herhangi bir büyük kromozomal yeniden düzenlemenin olmadığı basit bir evrimsel senaryoya işaret eder. İki komşu tetrad ise her biri aynı sadelik derecesindeki iki senaryodan biriyle açıklanabilir: tek bir 1R-öncesi kromozomdan türeyen bir 1R-sonrası kromozomu kırılmıştır (ayrışma) ya da her biri ayrı 1R-öncesi kromozomlardan türeyen iki 1R-sonrası kromozomu birleşmiştir (kaynaşma). Daha önce belirtildiği gibi, ikilenmemiş bir dış-grup tür, iki olası atasal konfigürasyon arasında ayrım yapmaya yardım edebilir.
Bununla birlikte, omurgalılara en yakın iki dış-gruptan ne gömlekliler (örn. Ciona grubu türleri) ne de başı-kordalılar (örn. batrak Branchiostoma floridae) bu amaca uygun değildir. İlki açık kromozom homolojileri belirlemek için fazla çeşitlidir; B. floridae genomu ise bilgi vermek için fazlasıyla parçaya ayrılmış durumdadır. Bu sorunu aşmak için daha önce yayımlanmış olan 17 Kordalı Bağlanım Grupları'nın (İng. Chordate Linkage Groups - CLG) yeniden yapılandırımını kullandık; bunlar aynı atasal kordalı kromozomundan türeyen insan genleri gruplarıdır. Yeniden yapılandırılmış bu proto-karyotip, 1R-2R olaylarından 50 milyon yıldan daha az bir zaman önce gelir ve Vertebrata ataya, varolan batrak genomundan çok daha kısa bir evrimsel mesafededir.
 FIG.4: Evrimsel senaryo modelleri.
FIG.4: Evrimsel senaryo modelleri.Dikkate değer şekilde, her bir Kordalı Bağlanım Grubu bir tane baskın BAB tetradla ilişkilendirilir. Bunun sonucunda, yedi komşu tetradın hepsi, 1R ile 2R TGİ'leri arasındaki kromozom kaynaşmalarından ortaya çıkar. Aslında, bir kromozom ayrışması, iki farklı tetrad üzerinden bir Kordalı Bağlanım Grubu'nun gen içeriğini ayırabilirdi. Yeri gelmişken, ayrıca kordalı ata ile 1R arasında bir kromozom kaynaşmasına ilişkin kanıt bulduk, çünkü 6 ve 7 numaralı Kordalı Bağlanım Grupları'ndan çıkan genlerin %75'ten fazlasının ardılları tek bir tetradda bulunuyor (tetrad 14). Bunun tersine, tetrad 13'ü bir Kordalı Bağlanım Grubu ile güvenilir biçimde ilişkilendiremiyoruz, bunun nedeni muhtemelen küçük bir tetrad olması. Dolayısıyla şu sonuca vardık ki 1R-öncesi karyotip 17 kromozomdan oluşuyor, ilk TGİ'den sonra 34 kromozoma ikileniyor ve ardından yedi kaynaşma geçiriyor. Ortaya çıkan 27 kromozom, ikinci TGİ'de ikilenerek, varolan yaklaşık 60.000 omurgalı türün kökenindeki 54 Vertebrata kromozomunu oluşturuyor.
2R'den sonraki kromozom evrimi
2R TGİ'den sonra farklı aşamalardaki ilave kromozom kaynaşmaları, bu karyotipin ardından geldi. Dört kaynaşma, Fig. 4c'de tanımlanan senaryoyu izledi; burada tek bir BAB iki tetradı birbirine bağlıyor (BAB 5, BAB 5a_152, BAB 3_22, BAB 10_240_2). Bunlar, hem ardıl hem de dış-grup genomlarına homolojileri nedeniyle, 2R TGİ ile Euteleostomi ata arasındaki döneme tarihlenebilir. Örneğin, BAB_10_240_2 tek bir tavuk kromozomuna (GG4), tek bir insan kromozomuna (X kromozomu) ve tek bir benekli zargana kromozomuna (LG7) homologtur, ki bunun da en sade şekilde uyduğu durum, bu BAB'ın zaten bu üç türün ortak atası olan Euteleostomi'deki tek bir kromozom olduğu durumdur. Beşinci bir kaynaşma, Euteleostomi ile Amniota arasındaki döneme tarihlenebilir: BAB 118 iki tetradda ortaktır ama tek bir tavuk kromozomu (GG1) ile homolog iken, benekli zargana genomunda gen sıralamasının aynılığındaki bozulma ve Japon pirinç balığı genomundaki DCS (İng. double conserved synteny ) örüntüsündeki bozulma, Amniota'ya giden soydaki bir kaynaşma ile tutarlıdır. Bu kaynaşmaları açıklayarak, 2R-sonrası Vertebrata'daki 54 kromozom, 50 kromozomlu (4 kaynaşma) bir Eueteleostomi karyotipine ve 49 kromozomlu (1 kaynaşma) bir Amniota karyotipine yol açmıştır. Özel bir Genomicus sunucusu (İng. server), bu çalışmada sunulan genomları karşılaştırmak ve çözümlemek için grafiksel bir arayüz sağlamaktadır.
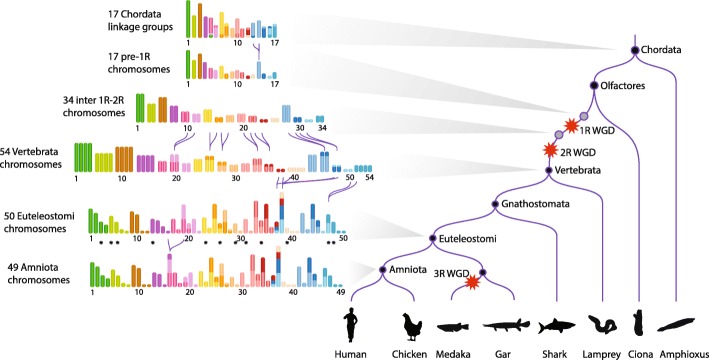 FIG.5: Kordalılardan Amniota'ya karyotiplerin yeniden yapılandırılmış evrimsel tarihi. FIG.5: Kordalılardan Amniota'ya karyotiplerin yeniden yapılandırılmış evrimsel tarihi.
FIG.5: Kordalılardan Amniota'ya karyotiplerin yeniden yapılandırılmış evrimsel tarihi. FIG.5: Kordalılardan Amniota'ya karyotiplerin yeniden yapılandırılmış evrimsel tarihi.1R-öncesi genom ile insan genomunun karşılaştırmalı genomiği
1R-öncesi omurgalı atası genomunun yeniden yapılandırılmışı ile varolan türler arasında karşılaştırmaya olanak tanımak için 17 1R-öncesi kromozomun her biri ile genler ilişkilendirdik. Amniota genleri arasında, sadece ohnologlar 1R-öncesi kromozomlarla güvenilir biçimde ilişkilendirilebilirdi çünkü ohnolog olmayan genler 1R TGİ'den sonra edinilmiş olabilirdi. Bu sorunu aşmak için muhafazakâr bir prosedür uygulayarak, 10.093 atasal Olfactores geninin 5052 tanesini, tahmin edilen 17 1R-öncesi kromozomla ilişkilendirdik. Olfactores ata, omurgalıların ve gömleklilerin ortak atası olup, Ensembl gen ağaçlarında yeniden yapılandırılan 1R-öncesi genomun kaynak yönünde en yakın atasıdır. Bu atasal genler kümesi, insandaki 8378 ardıl geni yoluyla insan genomuyla doğrudan bir bağlantı sağlar. İnsandaki bu ardılların her birini, kendi atasal 1R-öncesi kromozomunun rengiyle kimliklendirerek, 17 1R-öncesi kromozomun yapısının insan genomunda hâlâ çarpıcı şekilde belirgin olduğunu ve bazı kromozomların (örn. kromozom 14 ve 15) neredeyse bütünüyle tek bir 1R-öncesi kromozomdan gelen genlerden oluştuğunu gösterdik. İnsan kromozomları boyunca her 10 gende bir konumlandırılan 50 gen penceresinden, 1R-2R-sonrası ohnolog içeriğinin korunum derecesini ölçtük. Hox kümeleri A, B ve D'yi (ve daha az bir derecede Hox C'yi) çakıştıran üç bölge ön plana çıktı; bu da, bu ohnologların kümelenmesinin bilinen işlevsel önemiyle ilişkisine uygun.
 FIG.6: 17 kromozomdan oluşan 1R-öncesi karyotip ile insan karyotipi arası karşılaştırma.
FIG.6: 17 kromozomdan oluşan 1R-öncesi karyotip ile insan karyotipi arası karşılaştırma.Dört Hox kümesi 1R-öncesi kromozom 1'den gelmektedir ve biz de aynı ışık altında, tek bir 1R-öncesi lokustan geldiği ileri sürülen diğer paralog kümeleri inceledik. İnsanın 6.kromozomu üzerindeki MHC bölgesi, bağışıklık işlevleriyle ilgisiz çok sayıda gen içerir ama bunlar 1., 9. ve 19.kromozomlar üzerindeki 3 başka lokusta ohnologlara sahiptir. Bu 4 bölgenin hepsi 1R-öncesi 9.kromozomdan türemiştir. Benzer biçimde, FOX genleri kümesini içeren lokuslar karşılaştırılmış ve paraloglar paylaştıklar bulunmuştur ki bu da omurgalı evriminin erken dönemlerinde en-bloc (bütün hâlinde) ikilenmeleri akla getirir. Biz burada bunların 1R-öncesi 10.kromozomdaki benzersiz kökenini doğruladık. Tersine, insan genomundaki damgalanmış (İng. imprinted) genlerin farklı kümeleri için ortak bir köken bulunmamıştır (örn. 11.kromozom üzerindeki H19 lokusu, 6.kromozom üzerindeki IGF2R lokusu, 7.kromozom üzerindeki PON lokusu, 15.kromozom üzerindeki UBE3A lokusu); bu da bunların daha sonraları doğuran memelilerdeki bilinen dereceli belirişiyle uyumludur. O halde, atasal omurgalı kromozomları hakkında burada sunulan evrimsel senaryo, bu önemli gen ailelerinin evrimi konusundaki şu anki görüşümüzle uyum içindedir.
Son olarak, atasal Amniota ohnologlarının 1416 çiftinin (2 gen yitimi), 502 üçlüsünün (1 gen yitimi) ve 172 dörtlüsünün (yitim yok) insandaki ardıllarının Gene Ontology terimlerinin sıklığını analiz ettik ve çarpıcı bir örüntü bulduk: dörtlüler hem nöronal gelişimde hem de nöronal işlevde (sinaptik iletim) boldu; üçlüler kas gelişiminde (özellikle kalp) ve kas işlevinde (kasılma) boldu; çiftler (iki yitim) ise protein olgunlaşmasında ve organeller arası ulaştırmada boldu.
Tartışma
Ardından gelen 450 milyon yıllık süre zarfında 60.000'den fazla türe çeşitlenen omurgalıların atasına giden soydaki genlerin ve kromozomların evrimsel tarihini yeniden yapılandırmak için 61 hayvan genomunu çözümledik. Varolan genomları çözümleyen önceki çalışmaların tersine, kromozom ikilenmelerinin izini daha net belirlemek için biz önce dikkatli bir şekilde atasal Amniota genomunu yeniden yapılandırdık. Bunu yapma gerekçemiz, yeniden yapılandırılan atasal Amniota genomunun, ardından gelen 326 milyon yıllık evrim sırasında gerçekleşen sayısız yeniden düzenlemenin neden olduğu gürültüden yoksun olması gerektiğiydi. Gerçekten de bu yaklaşımın yararı, atasal Amniota genomundaki ohnologlar ile onların insan genomundaki ardıllarının dağılımları karşılaştırılarak, örneğin Hox kümelerini taşıyan kromozomlarda görülebilir. İkilenmiş kromozomlar arasındaki beklenen dört-taraflı ilişki, Amniota yeniden yapılandırımında çarpıcıdır ama insan genomunda bulanıktır. İstatistiksel güvenilirlik ve güç dolayısıyla yüksektir ve kromozom olaylarının daha önce mümkün olandan daha yüksek bir çözünürlüğüne olanak tanır. Böylece, bu yüksek sinyal-gürültü oranı, karmaşık istatistiksel adımlara veya algoritmik geliştirmelere gerek bırakmadan, ikilenme-öncesi genomlarının dolambaçsız bir yeniden yapılandırma stratejisine olanak tanır.
Nakatani ve ark. insan genomunu korunmuş omurgalı bağlanım gruplarına bölümleyerek bir omurgalı ikilenme-öncesi atasal genomunu yeniden yapılandırdığında, erken dönem omurgalı genomu evriminin anlaşılmasında ilerleme kaydedildi. Ohnologları tanımlamak için gömlekliler ve deniz kestanesi genleri kullanılarak, bunlar başka omurgalı, özellikle de Japon pirinç balığı genomları ile karşılaştırıldı. Bu strateji kullanılarak, yazarlar 1R-öncesi omurgalı atasının bir karyotipini çıkardı ama TGİ'ler-arası kromozom kaynaşmalarını veya ayrışmalarını çözüme kavuşturamadı ve 10 ilâ 13 tane 1R-öncesi kromozomun varolduğu sonucuna vardılar.
Burada, insan genomu yerine atasal Amniota genomu kullanarak, biz 1R-öncesi genom için 17 tane kromozom yeniden yapılandırımı gerçekleştirdik ve iki TGİ arasındaki 7 kaynaşmaya ilişkin kanıt bulduk; ayrışma için ise kanıt elde etmedik. Böylelikle yeni kromozomlar belirledik ve iki karyotip arasındaki başlıca farkları bulduk. Bizim çalışmamız, atasal omurgalı genomu evriminin geliştirilmiş bir resmini sunuyor; bunun nedeni biraz da yeniden yapılandırılmış Amniota genomuna dayalı olması ve ayrıca daha fazla türden edinilen bilgiye ve daha yeni tarihli genom notlandırmalarına dayanıyor olmasından ileri geliyor. Örneğin benekli zargana genomu, Japon pirinç balığı genomundaki gen sıralaması aynılığından daha bilgi vericiydi çünkü teleost TGİ'sinden önce gerçekleşmiş olan sayısız kromozom kaynaşmasından yoksundu.
Nakatani ve ark. ayrıca üç genomun (teleost TGİ-öncesi, tavuk ve omurgalı 1R-öncesi genomu) ikisi arasındaki korunuma dayanan bir "3'ün-2'si" kuralını kullanarak bir Osteichthyes 2R-sonrası genomunu da yeniden yapılandırmıştı ki o buradaki Euteleostomi ataya karşılık geliyor. Ancak burada, bizim Euteleostomi atamız, bu eski çalışmadan önemli ölçüde farklı; biz Euteleostomi genomunu 31 değil, 50 kromozomlu olarak tanımladık. Bu fark temelde, atasal teleosta giden soydaki kromozom kaynaşmalarının yüksek oranı nedeniyle karışıklık yaratan tavuk-teleost genom karşılaştırmalarından hatalı olarak çıkarsandığını düşündüğümüz kaynaşmalardan ileri geliyor. Aslında, atasal teleost balığın sadece 13 kromozom taşıdığı düşünülürse, burada atasal Euteleostomi için çıkarsanan 50 kromozom, teleostlara varan soydaki kromozom kaynaşmalarının oranının daha önce düşünülenden daha yoğun olması gerektiğine işaret eder; 100-150 milyon yılda 37 kaynaşma mertebesinde. Benzer biçimde, atasal eteneli karyotipi muhtemelen 23 çiftten oluşmaktadır ve 1R-2R tüm genom ikilenmelerinden sonra kromozom kaynaşması ile karyotip indirgemesinin tutarlı bir örüntüsüne işaret eder. Teleost ve memeli soylarındaki bu kaynaşmalar hem makro-kromozomları hem de mikro-kromozomları içerir ve muhtemelen bu soyların neden artık mikro-kromozomlar taşımadığını açıklar.
Burada tanımlanan atasal omurgalı kromozomlarının geliştirilmiş bir resmini kullanarak çeşitli soruları ele aldık. Öncelikle, ne Amniota genomunun yeniden yapılandırımında, ne de burada tanımlanan ohnolog çiftlerinin ortaya konmasında, omurgalı evriminin erken döneminde iki ardışık TGİ olduğuna ilişkin hiçbir varsayım yapılmadığına dikkat edilmelidir. Aslında, ohnologları seçmek için kullanılan ölçüt, sadece ikilenme tarihlerine ve yerel gen sıralaması aynılığına dayanarak, omurgalı genomu evriminin erken dönemlerinde bölümsel ikilenme, tek bir TGİ veya iki TGİ olması olasılıklarına açık kapı bırakmıştır. Ama bağlantı olarak ohnolog genleri kullandığımızda Amniota BAB'larının tetrad oluşturmak için hemen ilişkilendiği gerçeği, 1R-2R hipotezinin çarpıcı bir doğrulamasıdır. Bu senaryo üzerinde günümüzde aslında büyük ölçüde görüş birliğine varılmıştır ama bufa balığı (Petromyzon marinus) genom dizilimi ve tavuk genomu kullanılarak 1R-öncesi omurgalı genomunun yeniden yapılandırımının ardından, yakın zamanda yeniden tartışma başlamıştır. Bufa balığı bir Cyclostomata'dır; Cyclostomata ise Amniyotların ait olduğu Gnathostomata'nın kardeş bir grubudur ve her iki grup da ortak ata olarak 1R-öncesi genomu paylaşır. Bufa balığı genomu neredeyse 100 kromozomdan oluşur ve onun soyu, son TGİ'den kısa süre sonra Gnathostome'lardan ayrılmıştır. O hâlde muhtemelen gereğinden fazla gen kopyaları, her iki soyda da bağımsız olarak yitirilerek, eldeki bir Gnathostome (burada tavuk) kromozomunun, aynı atasal 1R-öncesi kromozomunda türeyen tüm bufa balığı kromozomlarıyla hâlâ homolojiler sergileme olasılığına açık kapı bırakır.
Smith ve ark.'ın bulguları, atasal 1R-öncesi kromozomlarının tavuk kromozomlarına oranının çoğunlukla 1:2 ve daha ender olarak 1:4 veya hatta 1:3 olduğunu ileri sürerek, her iki soyda atasal olan tek bir TGİ ile birlikte, en azından bufa balığı soyunda çok sayıda ilave bölümsel ikilenmeler olduğunu destekler. Ancak yukarıdaki çalışmayı tavuk yerine, yeniden yapılandırdığımız atasal Amniota genomu ile yinelediğimizde, net bir çoğunlukla 1:4 örüntüler belirdi; dolayısıyla iki ardışık TGİ'nin gerçekleştiğini destekledi. Bu ayrıca, böyle antik evrimsel olayları (günümüzden yaklaşık 450 milyon yıl önce) incelerken, aracı olarak atasal genom yeniden yapılandırımlarını kullanmanın yararını vurguluyor. Ek olarak, net 1:4 örüntüsün en sade açıklaması, Gnathostome ve bufa balığı soylarının ortak atasal tarihlerinde 1R-2R ikilenmeleri ortak olması ve Gnathostome'ların bufa balığı soyundan ayrılmasının 1R-2R ikilenmelerinden sonra gerçekleşmesi şeklinde oluyor.
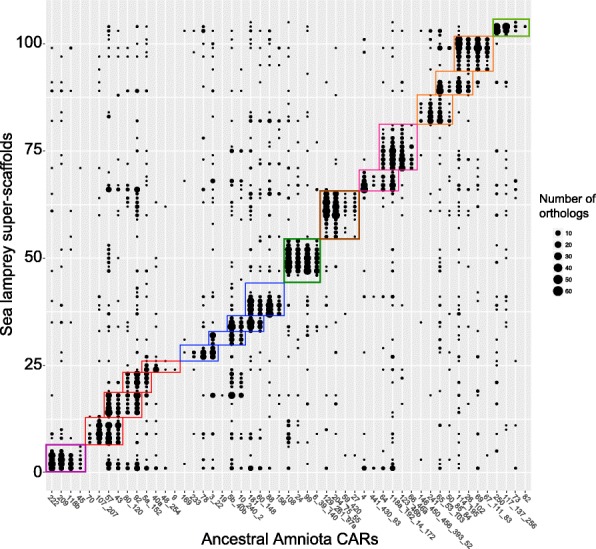 FIG.7: Atasal Amniota BAB'larının bufa balığı germ hattı genom düzenlemesinin süper-iskeleleri ile karşılaştırılması.
FIG.7: Atasal Amniota BAB'larının bufa balığı germ hattı genom düzenlemesinin süper-iskeleleri ile karşılaştırılması.Yeniden yapılandırılmış Amniota genomu tam değildir; 49 kromozom, BAB'larla ilişkilendirilen 15.854 atasal Amniota geninin sadece %80'ini içeriyor. 1R-öncesi kromozoma karşılık gelen tüm kromozom tetradları tam olmasına rağmen (yani 4 BAB'dan oluşuyor), 49 yeniden yapılandırılmış Amniota kromozomunun gen sayısı bakımından büyük farklar gösterdiği dikkatimizi çekti: en büyüğü 862 gen içerirken (kromozom 37), en küçüğü sadece 16 gen içeriyor (kromozom 49). Bu durum, yeniden yapılandırımlarında daha büyük güçlüklere yol açacak şekilde, daha az geni olan kromozomlar üzerindeki daha yoğun bir gen inaktivasyonu ve yitimi sürecini ya da bu kromozomlardaki daha yoğun oranda bir yeniden düzenlemeyi yansıtıyor olabilir. Az geni olan küçük kromozomlar, tetradlar arasındaki dağılımlarında, yeniden-diploidleşme (İng. rediploidization) sırasındaki gen silinmesine (delesyon) sistematik eğilime işaret edebilecek olan dikkat çekici herhangi bir örüntü izlemiyor. Bundan başka, eğer ilk TGİ'yi izleyen gen yitimi, iki ohnolog kromozom üzerinde, ikinci TGİ'nin sonucunda iki ikilenmiş kromozoma yayılmış olabilecek ayırt edici bir örüntü bıraktıysa olması bekleneceği gibi, kromozomların bir tetrad içinde çift yapılıp yapılamayacağına baktık. Burada yine öyle bir örüntü dikkatimizi çekmedi; bu da daha önce ileri sürüldüğü gibi iki TGİ'nin yakın aralıklarla gerçekleştiğine işaret ediyor olabilir; o nedenle gen silinmelerinin (diploidleşme) damgalarını bırakabilmek için fazla zamanı olmamıştır. Atasal genomlarının yeniden yapılandırımını daha iyi hâle getirmek ve 1R-2R tarafından bırakılmış daha silik örüntüleri belirleyebilmek için varolan amniyotların daha fazla yüksek-kalite genom dizilimleri gerekmektedir. Ancak, şu anki bu Amniota yeniden yapılandırımı ve onun ohnolog gen notlandırması, karmaşık filogenetik tarihler de dahil olmak üzere önemli soruların çözümlenmesine yardım edecek yeni çalışmalar için hâlihazırda sağlam bir temel sağlar.
Zargana genomunun çözümlenmesi sırasında, mikro-kromozomlar da dahil olmak üzere, zargana ile tavuk kromozomları arasında sık karşılaşılan bir 1:1 ilişkisini yazarlar fark ettiğinde ilginç bir soru ortaya atılmıştır. Küçük boyutları (tavukta genellikle 20 Mb altında), yüksek GC içeriği, yüksek rekombinasyon oranları ve yüksek gen yoğunluğu nedeniyle mikro-kromozomlar sıradışıdır. Zargana-tavuk karşılaştırması, mikro-kromozomların Euteleostomi'deki atasal özellikler olduğuna işaret eder ki bu da onların 1R-2R ikilenmeleri yoluyla olan kökeni sorusunu gündeme getirir. Oniki tane zargana ve tavuk mikro-kromozomu homologtur ve sadelikli bir şekilde Euteleostomi'de atasal olarak bulundukları düşünülebilir. Bunlar 7 ilâ 11 tetrada dağılmıştır (bazı Euteleostomi mikro-kromozomları iki tetraddan türemiştir), bunların her biri hem mikro-kromozomlar hem de makro-kromozomlar içerir. Bu durum onların oluşum zamanı sorusunu yanıtsız bırakır: 1R'den önce, 1R ile 2R arasında veya 2R'den hemen sonra.
Burada tanımlanan 2R-öncesi karyotip ve onun evrimi, varolan omurgalı genomlarında 1R-2R'nin etkisini incelemek için bir çerçeve sağlar; özellikle de çözümlemenin temelini oluşturan filogenetik açıdan tutarlı ohnolog aileleri kümesi yoluyla. Örneğin, ohnologların olası bir taraflı muhafazasını açıklamak için daha önceki çalışmalar alternatif hipotezler sunmuştur; komplekslerdeki proteinlerin stokiyometrik (tam oranlı) dengesi için bir gereklilik veya genomdan silindikleri takdirde uyumluluk üzerinde baskın olumsız sonuçları sayesinde hastalıklarla ilgisi olan genlerin tercihli olarak muhafazası da dahil. Ohnologların ayrıca gelişimsel genlerde ve düzenleyici işlevde bol bulundukları gösterilmiştir. Taraflı ohnolog muhafazasını açıklayacak tam bir mekanizma henüz ortaya konmamıştır. Bunun nedeni, bu çalışmaların varolan gen kopyalarına dayanması ve 1R-2R TGİ'lerinden beri 500 milyon yıllık evrimin geçirilmiş olması olabilir. Ohnolog Amniota genlerinin insandaki ardıllarını kullanarak, iki gen yitimine uğrayan, bir gen yitimine uğrayan veya bir gen dörtlüsü olarak kalan ohnolog ailelerinde, biyolojik işlevlerin çarpıcı bir örüntüsünün tercihli olarak muhafaza edildiğini gösterdik. Üç çeşit ohnolog ailesi arasında işlevlerin böylesine ayırt edici ve özel bir dağılımının olması, ohnolog korunumu ve yitiminin erken dönem oranlarını yöneten güçlü işlevsel kısıtlar sorusunu gündeme getirir. Bu örüntülerin, omurgalı evriminin erken dönemlerinde, genlerdeki gereksiz fazlalığın maksimum olduğu ve evrimsel yenilikler için bir tuval sağladığı zamanda, artırılmış organizmal karmaşıklığa yol açacak şekilde, seçilimle en çok hedeflenen işlevleri ve anatomik parçaları (merkezi sinir sistemi ve kaslar) yansıtıyor olması mümkündür. Bu ayrıca yeniden yapılandırılmış atasal genomlara dayalı olarak yapılan karşılaştırmalı genomiğin yararlarını göstermektedir.
Sonuç
Biyoloji tarihsel bir bilimdir, ama bu tarihsel boyutun doğrulanması çoğu zaman güçtür çünkü atasal durumları belgelemesi gereken genomik kayıtlar eksiktir. Uygulamada, bu bilgi yoksunluğu, yaşayan farklı organizmalar üzerinden varılmış sonuçları birleştime becerimize ve karşılaştırmalı genomiğin tüm yararını görmemize köstek olur. Burada, omurgalı genomunun erken dönem tarihinin ayrıntılı olarak yeniden yapılandırılması, yaşayan memelilerin, kuşların, sürüngenlerin, ikiyaşayışlıların ve balıkların kökeninde temel teşkil eden olayların daha iyi anlaşılması yolunda bir adımdır. Varolan bu hayvanların evriminin incelenmesi için ortak bir referanstan yeni bir perspektif sağlar.
Kaynak ve İleri Okuma
- Chromosome evolution at the origin of the ancestral vertebrate genome https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6193309/
- OHNOLOGS http://ohnologs.curie.fr/
Etiket
Projelerimizde bize destek olmak ister misiniz?
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek Ol
Yorum Yap (0)
Bunlar da İlginizi Çekebilir
19 Ağustos 2019
Sperm Cinsiyetini Belirlemenin Kolay Bir Yolu Keşfedildi

14 Nisan 2018
Y Kromozomu Yok Oluyor. Peki Erkeklere Ne Olacak?
30 Haziran 2019
Memelilerde Dozaj Telafisi
27 Haziran 2019
Heterokromatin Kavramına Genel Bir Bakış
26 Kasım 2016
Bir Kromozomun Yalnızca Yarısını DNA Oluşturuyor

12 Ekim 2017
İnsan Genomu Kendini Katlarsa
27 Haziran 2019
Kromozom Haritaları: Idiogramlar
Bağış Yap, Destek Ol!
Projelerimizde bize destek olmak istersen
Patreon üzerinden aylık veya tek seferlik
bağışta bulunabilirsin.
En Çok Okunan
Bu Ay Öne Çıkanlar



2
İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

Gürkan Akçay
Boğaziçi Üniversitesi - Yazar / Editör
E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.