
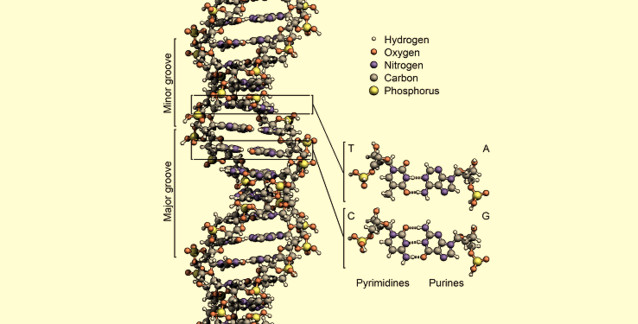

Deoksiribonükleik asit (DNA), canlı organizmaların çoğunun genetik bilgisini taşıyan moleküldür; söz konusu çoğunluğun dışında kalanlarda ise genetik bilgi ribonükleik asit (RNA) molekülü tarafından taşınır. DNA, hücrelerde nesilden nesile kopyalanabilir (DNA eşlenmesi); proteinlere çevrilebilir (DNA'nın RNA'ya yazımı ve sonra proteinlere çevrimi); gerektiğinde onarılabilir (DNA onarımı).
DNA, nükleotid (veya mono-nükleotid) adı verilen birimlerden oluşan bir polimerdir. Nükleotidlerin başka işlevleri de vardır: Enerji taşımada ATP, GTP; hücresel solunumda NAD, FAD; sinyal aktarımında çevrimsel AMP; koenzim olarak CoA, UDP; vitamin olarak nikotinamid mononükleotid, B2.
Protein terimlerini kullanarak, molekülün birincil, ikincil, üçüncül ve dördüncül yapılarından söz edebiliriz:
1. Molekülün Birincil Yapısı: Kovalent Omurga ve Yanlarda Bazlar
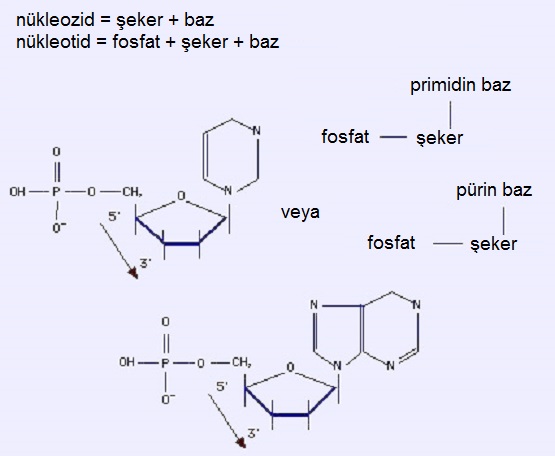
Bir nükleozid, bir şeker + bir azotlu bazdan oluşur.Bir nükleotid, bir fosfat + bir şeker + bir azotlu bazdan oluşur.DNA'da nükleotid bir deoksiribonükleotiddir (RNA'da nükleotid ise bir ribonükleotiddir.)
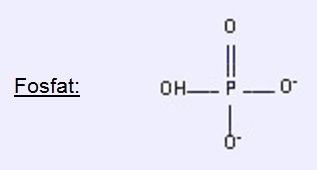
1.1. Fosforik Asit
Bir fosfat grubu verir.
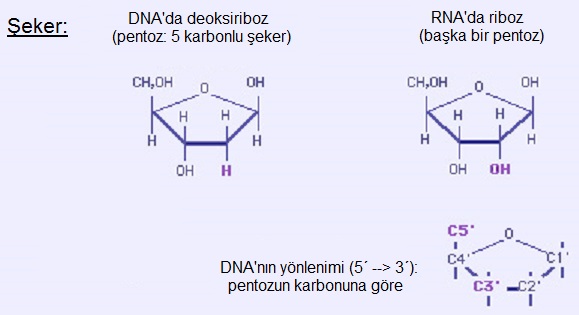
1.2. Şeker
Halkasal bir pentoz (5 karbonlu şeker) olan deoksiriboz. (RNA'daki şeker ribozdur.) Şekerdeki karbonlar 1' den 5' ye kaydedilir. Azotlu bazlardan bir azot atomu C1' ye tutunarak (glikozidik bağ), fosfat ise C5' ye tutunarak (ester bağ) nükleotidi oluşturur. Dolayısıyla nükleotid şöyledir: fosfat - C5' şeker C1' - baz.
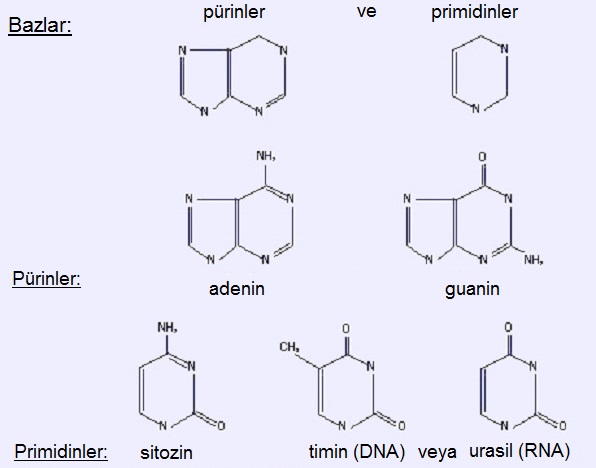
1.3. Azotlu Bazlar
Aromatik hetero-halkalılar (bir bileşiğin aromatik olması için halkalı yapıda olması, halkanın düzlemsel olması, halkada konjuge çift bağların bulunması ve halkanın, (4n+2) sayıda π-elektronu taşıması (Hückel Kuralı) gerekir.); pürinler (adenin & guanin) ve primidinler (sitozin & timin; RNA'da timin yerine urasil bulunur) vardır. Başka azotlu bazlar da mevcuttur; özellikle belirtmek gerekirse, yukarıda söz edilenlerden türeyen metillenmiş bazlar vardır. Bazların metillenmesinin işlevsel bir rolü bulunur.
Terimler:
Nükleozid adları:
- DNA'daki deoksiribonükleozidler: deoksiadenozin, deoksiguanozin, deoksisitidin, deoksitimidin.
- RNA'daki rinonüklozidler: adenozin, guanozin, sitidin, uridin.
Nükleotid adları:
- DNA'daki deoksiribonükleotidler: deoksiadenilik asit, deoksiguanilik asit, deoksisitidilik asit, deoksitimidilik asit.
- RNA'daki ribonükleotidler: adenilik asit, guanilik asit, sitidilik asit, uridilik asit.
2. Molekülün İkincil ve Üçüncül Yapıları: DNA'nın Üç Boyutlu Şekli (Konformasyonu)
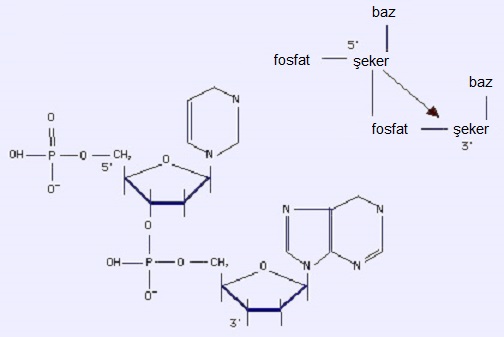
2.1. Dinükleotidler
Dinükleotidler, iki mononükleotid arasındaki fosfodiester bağıyla oluşur. Bir mononükleotidin fosfatı (şekerinin C5´ sünde), bir önceki mononükleotidin şekerinin C3´ süyle bağlantılı olur. O hâlde bir fosfatla başlıyoruz, bir 5' şeker (+baz) ve o şekerin 3' sü var ki o da ikinci bir fosfat-5' şekere bağlı. Dolayısıyla bağ ve molekülün yönlenimi 5' --> 3' olur. Polinükleotidler, monomerlerin genel bir 5' --> 3' konfigürasyonuyla ardışık eklenmelerinden meydana gelir. Molekülün omurgası, fosfat-şekerlerin kovalent bağlanarak arka arkaya gelmesiyle ve yanlarda bazların yer almasıyla oluşur.
2.2. DNA Molekülü
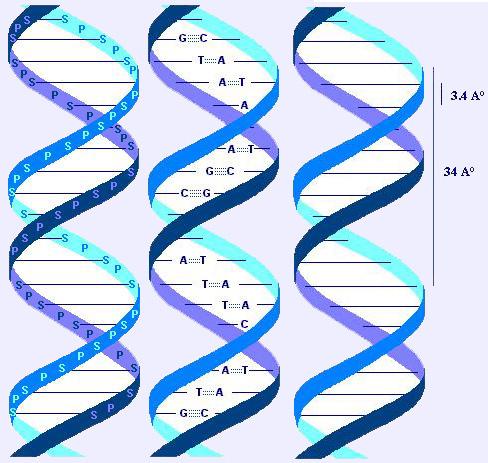
DNA, iki tane saat yönünde kıvrılan helezonik zincirden (iplikçikten) oluşur. Bu iplikçikler, çapı 20A° olan bir ikili helezon (ikili sarmal) meydana getirecek şekilde bir eksen etrafında bukleleşir. İplikçikler antiparaleldir (yani bunların 5' --> 3' yönlenimleri karşıt yönlüdür). Polimerin genel görünümü, iki baz arası uzaklığa karşılık gelen 3.4 A° lük bir periyodiklik sergiler. Ayrıca 34 A° lük bir periyodiklik daha vardır ki o da bir helezon bükümüne (ve ayrıca 10 baz ikilisine) karşılık gelir.
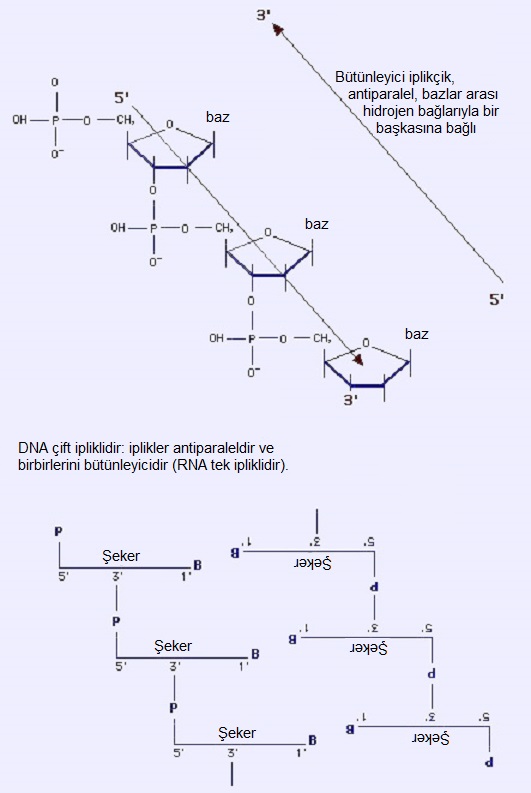
2.2.1. Hidrojen Bağları: Bazlar Eşleşiyor
Bazlar (hidrofobik) iç kısımda bulunur; düzlemleri ikili sarmalın eksenine diktir. Dış kısım (fosfat ve şeker) hidrofiliktir. Bir iplikçiğin bazları ile diğerininkiler arasındaki hidrojen bağları, iki iplikçiği birarada tutar (çizimde kesikli çizgi). Bir iplikçikteki bir pürin, diğer iplikçikteki bir primidine bağlanmalıdır. Sonuç olarak, pürin artıklarının sayısı, primidin artıklarının sayısına eşittir.
A ile T bağlanır (2 hidrojen bağıyla).G ile C bağlanır (3 hidrojen bağıyla: daha kararlı bağ: 5.5 kcal X 3.5 kcal).
Not: DNA'daki A miktarı T miktarına, G miktarı da C miktarına eşittir. Bu kaşılıklılık (A<->T ve G<->C) iki iplikçiği bütünleyici yapar. Birbirlerinin şablonudurlar ve bu özellik birebir eşlenmeyi sağlar (yarı-korunumlu eşlenme: şablon görevi gören bir iplikçik korunur; onun karşısına uygun yenisini sentezlenir.)
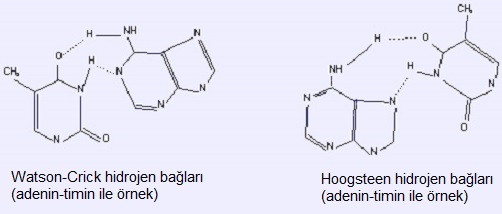
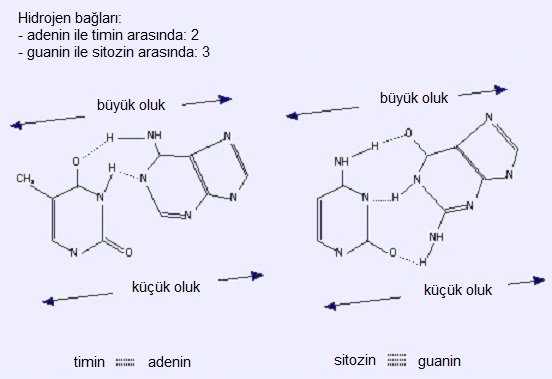 Not: Baz eşleşmesindeki hidrojen bağları bazen yukarıda tanımlanan Watson&Crick modelinden farklı olur; pürinin N1 atomu yerine N7 atomu kullanılır (Hoogsteen modeli).
Not: Baz eşleşmesindeki hidrojen bağları bazen yukarıda tanımlanan Watson&Crick modelinden farklı olur; pürinin N1 atomu yerine N7 atomu kullanılır (Hoogsteen modeli).
2.2.2. Büyük Oluk ve Küçük Oluk
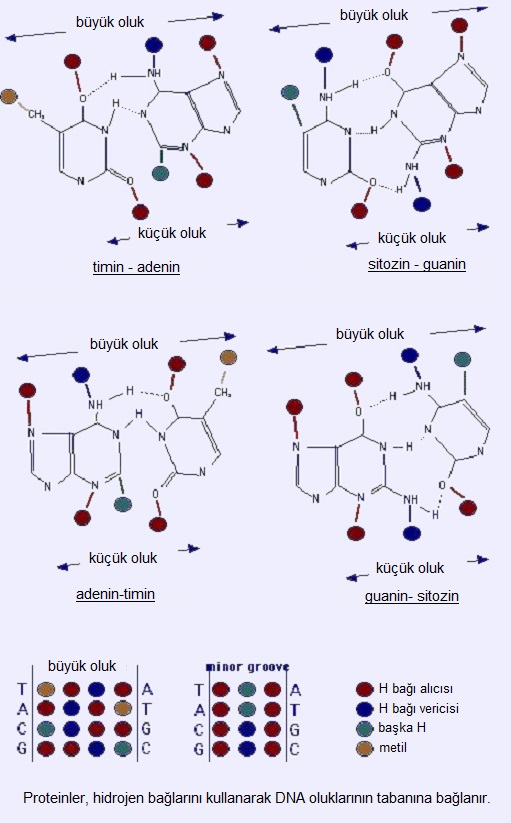
İkili sarmal kaskatı ve akmaz bir molekül olup, muazzam uzunlukta ve küçük genişliktedir. Bir büyük oluğu, bir de küçük oluğu bulunur. Büyük oluk derin ve geniştir; küçük oluk ise dar ve sığdır.
DNA-protein etkileşimleri, hücre yaşamında temel süreçlerdir (yazım etkinleştirilmesi ve baskılanması, DNA eşlenmesi ve onarımı). Proteinler, spesifik bağlarla (hidrojen bağları) ve spesifik olmayan bağlarla (van der Waals etkileşimler, genelleştirilmiş elektrostatik etkileşimler) DNA oluklarının tabanına bağlanır. Proteinler H-bağı vericilerini, H-bağı alıcılarını, metil gruplarını (hidrofobik) tanır; sonuncusu özel olarak büyük oluktadır. Büyük oluk için 4 olası tanıma şablonu varken, küçük oluk için 2 tane vardır. Bazı proteinler DNA'ya büyük oluğundan, bazıları küçük oluğundan bağlanır; bazılarının ise ikisine birden bağlanması gerekir.
 Notlar:- İki iplikçiğe "artı" ve "eksi" iplikçikler veya "doğrudan" ve "ters" iplikçikler denir. İki iplikçikten birinin kodlayıcı dizilimi taşıdığı belirli bir konumda, diğer iplikçiğin de kodlayıcı dizilimler taşıması (olanaksız olmasa da) pek muhtemel değildir.- DNA in vivo (canlı içinde) iyonlaştırılır ve bir poli-anyon gibi davranır.
Notlar:- İki iplikçiğe "artı" ve "eksi" iplikçikler veya "doğrudan" ve "ters" iplikçikler denir. İki iplikçikten birinin kodlayıcı dizilimi taşıdığı belirli bir konumda, diğer iplikçiğin de kodlayıcı dizilimler taşıması (olanaksız olmasa da) pek muhtemel değildir.- DNA in vivo (canlı içinde) iyonlaştırılır ve bir poli-anyon gibi davranır.
Yukarıda tanımlanan şekildeki ikili sarmal, DNA'nın "B" formudur; in vivo en sık rastlanan formdur ama in vivo ve in vitro (deney tüpü içinde, yani canlı dışında) başka formlar da mevcuttur. "A" formu B-DNA'ya benzer ama ondan daha az hidratlıdır ve in vivo bulunmaz.
2.3. B-olmayan DNA
DNA hareket eden, kımıldanan, jimnastik yapıp dans eden bir moleküldür. Aşağıda sözü edilen yapıların işlevsel rollerinin olduğu kanıtlanmıştır; öte yandan, DNA kırılmalarına ve silinmelerine, güçlendirmeye, rekombinasyona ve mutasyonlara çanak tutabilirler.
Terimler:Palindrom: Tersten okunduğunda, düz okunuşuyla aynı olan sözcüklerdir (örn. "DNA LAND"). DNA sık sık palindromlarla oynar.
2.3.1. Z-DNA
- Z formu, omurganın zigzag koformasyonlu olduğu (B-DNA'dan daha pürüzlüdür), solak bir ikili helezondur. Sadece bir oluk gözlemlenmiştir ve küçük oluğu anımsatır; baz çiftleri yanlarda, eksenden uzaktadır. Bazlar (B-DNA'da eksene yakın büyük oluğu oluştururlar) burada dış yüzeydedir. Fosfatlar birbirlerine B-DNA'da olduğundan daha yakınlardır. Z-DNA nükleozom oluşturamaz.
- Yüksek bir G-C içeriği Z konformasyonuna çanak tutar. Sitozin metillenmesi ve in vivo bulunabilen spermin ve spermidin gibi moleküller Z konformasyonunu dengeleyebilir.
- DNA dizilimleri B formu ile Z formu arasında değişebilir: Z-DNA in vivo bir geçiş formudur.
- Z-DNA oluşumu genlerin yazımı sırasında, aktif olarak yazımı yapılan genlerin başlatıcılarına yakın olan yazım balatma bölgelerinde gerçekleşir. Yazım sırasında, RNA polimerazın hareketi, yazım bölgesinin aşağısına pozitif süper-sarım ve yukarısına negatif süper-sarım tetikler. Yukarıya negatif süper-sarım Z-DNA oluşumuna çanak tutar; Z-DNA işlevlerinden biri negatif süper-sarımı soğurmak olabilir. Yazımın sonunda, topoizomeraz DNA'yı B konformasyonuna geri salar.
- Bazı proteinler Z-DNA'ya bağlanır; özellikle belirtmek gerekirse, çift-iplikli RNA adenozin deaminaz (ADAR1) adlı Z-DNA'ya bağlanan çekirdeksel-DNA-düzenleyici enzim bağlanır. Bu enzim pre-mRNA'da adenini inozine dönüştürür. Ardından, ribozomlar inozini guanin olarak yorumlar ve bu epigenetik modifikasyonla kodlanan protein farklı olur.
- Z-DNA antikorlar, lupus eritematozus hastalığı ve diğer otoimmün hastalıklarda bulunur.
- Çift iplikli RNA (dsRNA), bir Z-konformasyonuna girebilir.
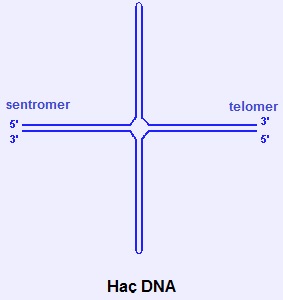
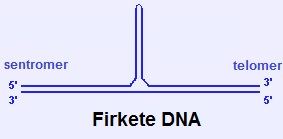
2.3.2. Haç DNA ve Firkete DNA
- Holliday bağlantıları (rekombinasyon sırasında oluşur), haç biçimli yapılardır. Polipürin/poliprimidin DNA ipliklerinin ters dönmüş (veya ayna) yinelemeleri (palindromları) de, iplik-içi çiftlenme yoluyla haç biçimli veya firkete biçimli yapılar oluşturabilir.
- Translokasyon t(11;22)(q23;q11) kesim noktalarında palindromik AT zengini yinelemeler bulunur; bu bilinen tek yinelenen yapısal karşılıklı translokasyondur.
- Rekombinasyondan sonra nükleazlar Holliday bağlantılarına bağlanır ve yarar. HMG proteinleri ve MLL gibi başka iyi bilinen proteinler de haç DNA'ya bağlanabilir.
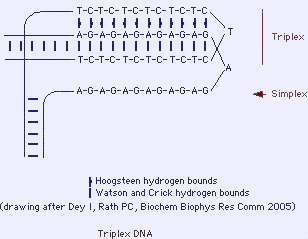
 2.3.3. H-DNA veya üç-katlı DNA
2.3.3. H-DNA veya üç-katlı DNA
- Polipürin/poliprimidin DNA ipliklerinin ters dönmüş yinelemeleri (palindromları), üç katlı yapılar (üçlü helezon) oluşturabilir. Bir üç-iplikli artı bir tek iplikli DNA oluşur.
- H-DNA gen ifadelenmesinin düzenlenmesinde ve RNA'lar üzerinde işlevsel bir rol alabilir (örn. yazım baskılanması).
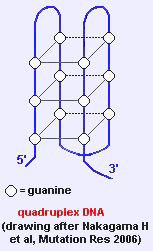
 2.3.4. G4-DNA
2.3.4. G4-DNA
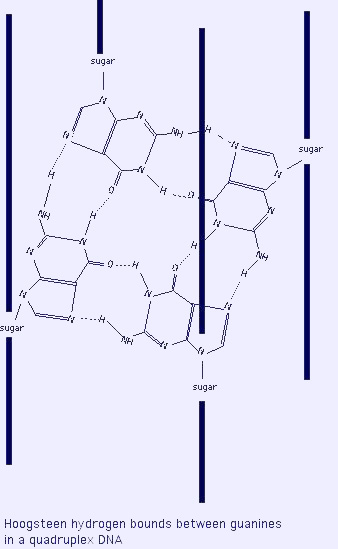
- G4-DNA veya dört-katlı DNA: Çift iplikli GC zengini dizilimin kendi üzerine katlanması, 4 guanin arasında ("G4") Hoogsteen baz çiftlenmesi oluşturur; oldukça kararlı bir yapıdır. Sıklıkla genlerin başlatıcılarının yakınında ve telomerlerde bulunurlar.
- Mayoz ve rekonbinasyondaki rolü, düzenleyicilik olabilir.- RecQ ailesi helikazları G4-DNA'yı çözebilir (örn. BLM, Bloom sendromundaki mutant gen).
3. Molekülün Dördüncül Yapısı: Kromatin
DNA proteinlerle (histonlar ve histon olmayan proteinler) birlikte kromatini oluşturur. DNA, bir bütün olarak asidik (negatif yüklü) olup, histon adı verilen bazik (pozitif yüklü) proteinlere bağlanır. İnsan haploid genomunda 3 x 10 9 nükleotid çifti vardır; bunlar, 23 kromozoma dağılmış durumdaki 30.000 civarında geni temsil eder.
4. Diğer Konular
4.1. DNA ve Mitokondriler
- DNA hücrelerin çekirdeğinde bulunur ama mitokondrilerde de küçük bir miktar DNA vardır.-
- Mitokondrilerin, ökaryotik hücrelerle içten-ortakyaşam (endo-simbiyoz) ilişkisine giren arkeobakterilerden türediği düşünülmektedir.- Mitokondrilerin genetik kodu, "evrensel" denilen koddan biraz farklıdır (evrensel kodda UGA, AUA, AGA, AGG sırasıyla DUR, Ile, Arg, Arg anlamında iken, memeli mitokondrilerinde yine sırasıyla Trp, Met, DUR, DUR anlamına gelir; başka türlerin mitokondrilerinde ise daha başka anlamlara gelebildiği görülmüştür).- Belli bir mitokondrideki DNA kopyalarının sayısı değişkendir.
- Mitokondriyal DNA daireseldir, bir ağır bir de hafif zinciri olur; intronları yoktur ve kodlamayan herhangi bir dizilimi yoktur.
- Mitokondrilerdeki genler elektron taşınımı, ribozomik RNA'lar (rRNA) ve aktarım RNA'ları (tRNA) ile ilgilidir.
- Her bir DNA ipliğinin yazımı yapılır, sonra mRNA'lara ayrılır fakat ayrıca rRNA'lara ve tRNA'lara da ayrılır.
Not: Mitokondriler, hücrenin sitoplazmasından alınan (ve çekirdek tarafından kodlanan) proteinleri de kullanır; şimdiye dek, apoptoz vakası dışında mitokondriden proteinlerin sitoplazmaya salınmasına rastlanmamıştır.
4.2. DNA Denaturasyonu
İkili sarmal, in vitro ısı, aşırı ph ve başka koşullarla (üre,...) çözülüş geçirebilir. Bir erime noktası hesaplanabilir; bu, A/T'de 2 hidrojen bağı varken G/C'de 3 olup daha kararlı bağlandığı gerçeğine bağlı olarak, incelenen örneğin A/T'ye karşı G/C oranının karakteristiğidir.
Denaturasyondan sonra DNA'nın fiziksel özellikleri değişir; örn. hiperkromik etki: 260 nm'de ışık emilimi denature DNA'da ikili helezon DNA'dan yüksektir. Işık emilimi A/T'ye karşı G/C oranına göre de değişir: A/T zengini örneklerde daha yüksektir.
DNA denaturasyonu,1- A/T'ye karşı G/C içeriğinin ölçülmesine olanak tanır;2- in situ (hücre içinde) hibritleştirme tekniklerinin temelini oluşturur.- DNA: molecular structure http://atlasgeneticsoncology.org/Educ/DNAEngID30001ES.html
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek OlBunlar da İlginizi Çekebilir







En Çok Okunan
Bu Ay Öne Çıkanlar



İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.