
 Porter K/Getty Images
Porter K/Getty Images

Öz
Ökaryotların evrimi ve çeşitlenmesi, mikroorganizmalar tarafından sürekli olarak kolonize edilmeleri sayesinde gerçekleşmiştir. Bünyelerine yerleşen mikroorganizmaların kimileri, ökaryotlara yeni metabolik beceriler kazandırarak, konaklarının ekolojik yayılımlarını yükseltmiş ve sıklıkla da evrimsel çeşitlenmelere yol açmıştır. Bazı mikroorganizmalar konaklarına fotosentez veya selüloz parçalama gibi birincil metabolik yolaklar bahşederken, bazıları da ikincil metabolizma repertuarını genişletmiştir; doğal düşmanlara karşı koruma sağlayan toksinlerin sentezlenmesi gibi. Mikroorganizmaların konağın sıhhatine katkıda bulunmasının bir diğer yolu da büyümeyi, gelişimi, davranışı ve diğer işlevleri düzenleyen ökaryotik-sinyalleme ağlarını modüle etmeleri ile gerçekleşir. Yarattıkları etkinin konağa yararlı etkileşimlere dayalı olması gerekmez; mikrobik ürünlerin ipuçları olarak ökaryotik kullanımının ya da konak-mikrop çekişmesinin bir sonucu olabilir. Bu yollarla, ökaryot-mikrop etkileşimleri, ökaryotların evrimsel çeşitlenmesinde ve işlevlerinde ayrılmaz bir rol oynar.
Ökaryotlar yalnız yaşamaz. Görünürde hiçbir hastalık etkisi olmadan, yüzeylerinde ve içlerinde canlı bakteri hücreleri (öbakteriler ve arkeler) ile çoğu zaman ökaryotik mikroorganizmalar da taşırlar. Ayrıca, varolan tüm ökaryotların, Rickettsiales grubuna dahil olan ve sonraları mitokondrilere evrilen hücre-içi bakterilerin ortaklığından türediğine ilişkin ikna edici kanıtlar vardır (Williams et al. 2007); bu da kalıcı ortaklıklar kurma eğiliminin çok eski evrimsel kökleri olduğuna işaret eder. Bu açıdan ökaryotlar, bakterilerden farklıdır; bakterilerin sadece bir kısmı (özellikle belirtmek gerekirse, öbakterilerin tahminen 52 şubesinin yaklaşık 11 tanesinin üyeleri (Sachs et al. 2011) ile arkelerin ufak bir azınlığı (Gill and Brinkman 2011)) ökaryotlarla ortaklık kurar.
Ökaryotlarla ilişkilendirilen mikrobiyotaya şu an gösterilen ilgi, mikrobiyal toplulukların kültürden-bağımsız analizindeki önemli teknolojik ilerlemelerle (özellikle de mikroorganizmaları belirlemek ve saymak için kullanılabilen yüksek çıktılı dizileme yöntemleriyle) başladı (Caporaso et al. 2011; Zaneveld et al. 2011). İnsan Mikrobiyomu Projesi (commonfund.nih.gov), MetaHIT (metahit.eu) ve diğer girişimler, insanlarla, diğer hayvanlarla, bitkilerle, mantarlarla, tek-hücreli ökaryotlarla (protistlerle) ve ayrıca abiyotik habitatlarla ilişkilendirilen mikroorganizmaların işlevsel becerileri ve taksonomik çeşitliliği hakkında emsali görülmemiş bilgiler veriyor (Qin et al. 2010; Muegge et al. 2011; Human Microbiome Project 2012a; Lundberg et al. 2012; Bourne et al. 2013). Yapılan araştırmaların çoğu öbakterilere odaklanmaktaydı ama mikrobiyotanın ökaryotik üyeleri, özellikle de mantargiller konusundaki araştırmalar artıyor (Iliev et al. 2012; Findley et al. 2013).
Teknolojik değişimle yönlendirilmiş olsa da, kültürden-bağımsız şekilde yapılan bu insan mikrobiyotasının ve diğer ökaryotların mikrobiyotalarının incelemeleri, bizim kavramsal anlayışımız üzerinde derin sonuçlar doğuruyor. Özellikle belirtmek gerekirse, 20.yy boyunca besin üretiminin ve halk sağlığının iyileştirilmesinde kilit rol oynamış olan "hastalıkların mikrop kuramı"nın, hayvanlarla ve bitkilerle ilişkilendirilen tüm mikroorganizmaların patojen (hastalık yapıcı) olduğu şeklindeki yaygın ama hatalı inanca da yol açtığının farkına varanlar artıyor. Bu modası geçmiş algının yerini, ökaryotların kronik olarak iyi huylu ve yararlı mikroorganizmalarla enfekte hâlde olduklarının ve mikrobiyota etkinliği veya bileşimi üzerindeki bir bozukluk sonucunda hastalık görüldüğü kabûlü alıyor (McFall-Ngai et al. 2013; Stecher et al. 2013).
Bu makale, mikroorganizmalarla ortakyaşamın (simbiyoz), ökaryotik konakların özellikleri üzerindeki egemen etkilerini ve ökaryotların evrimsel tarihinde meydana getirdiği sonuçları özetliyor. Ortaklıkların büyük bir çoğunluğu için ortakyaşamın etkileri iki çeşit etkileşime atfedilebilir. İlk etkileşim -"yeni becerilerin kaynağı olarak ortakyaşam"- ökaryotik konakta olmayıp, mikrobiyal ortakta bulunan metabolik veya başka özellikler üzerine kuruludur. Bu becerilere erişim kazanan ökaryotlar daha iyi besin, doğal düşmanlara karşı koruma veya başka seçimsel önem taşıyan özellikleri çeşitli kereler türetmiştir. İkinci etkileşim -"sağlığın ortakyaşamsal temeli"- büyüme hızı, bağışıklık işlevi, besin dağıtımı ve davranış dahil olmak üzere çok sayıda özelliğin mikrobiyal modülasyonu yoluyla ökaryot konağın kazandığı sıhhat ve dinçliği kapsar; etkiler, konakta olmayan belirli mikrobiyal becerilere atfedilemese bile. Ortakyaşamın sağlık açısından yararlarının çoğu zaman, ökaryot konakların gelişimsel ve fizyolojik işlevlerinin koordine edildiği sinyalleme ağlarının mikrobiyal modülasyonunun bir sonucu olduğuna ilişkin kanıtlar artmaktadır.
Bu makale üç bölümden oluşuyor: Hayvanlarda, bitkilerde ve diğer ökaryotlardaki bir dizi ortakyaşarlıktan belirli örneklerle aydınlatılan önemli süreçler ve şablonlarla iki etkileşim çeşidi sırayla ele alınıyor; daha sonra sonuç yorumlarında, ökayotlarda ortakyaşam hakkında yanıtlanmamış bazı önemli sorulara değiniliyor. Bu makale, ökaryotik evrimde ortakyaşamın genel ilkelerine ilişkin sözü edilen ortaklıkların tüm çeşitliliğini özetlememektedir; ilgili okurlar için Douglas (2010) önerilir.
Yeni Becerilerin Kaynağı Olarak Ortakyaşam
Metabolizma Üzerine Kurulu Ortakyaşamların Kadim Evrimsel Kökleri
Ökaryotlar, kendilerinde olmayan işlevlere sahip başka organizmalarla ortakyaşam kurarak, karmaşık metabolik becerilere çeşitli kereler erişim kazanmıştır. Organizmayı (sadece genleri değil) elde ederek, ökaryotlar belirli enzimleri kodlayan genleri kazanmanın yanı sıra, onların düzenlenmiş ifadesi için olan genetik ve hücresel mekanizmayı da kazandı. Ortakyaşamla metabolik beceriler edinme kapasitesinin, varolan tüm ökaryotların ortak atasında olduğu bellidir ve ökaryotları doğuran soydaki metabolik sınırlandırmalarla bağlantılandırılabilir. Özellikle belirtmek gerekirse, bu soyun ototrofik karbon bağlama ve aerobik solunum kapasitesi bulunmaz; bu özellikler öbakterilerde (tahminen iki grubun ortak atasından daha yakın zamanda) evrilmiştir.
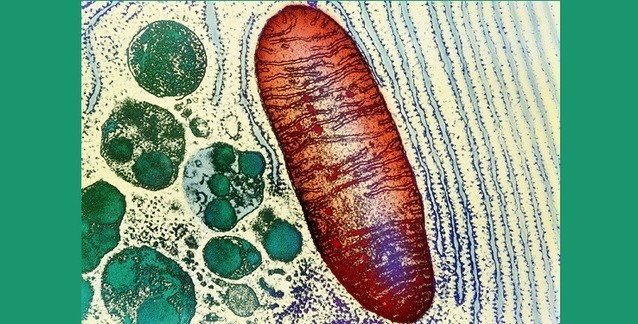
Mitokondrilerin α-proteobakteriyel atası ile ortaklık muhtemelen 1-2 milyar yıl önce evrilmiştir. Varolan tüm ökaryotlar ya mitokondrilere sahiptir ya da mitokondrisel atalardan türemiştir ve bu durum, mitokondrilerin ediniminin ökaryotların evrimsel kökenini tanımlama olasılığını yükseltir (Embley and Martin 2006). (Yine de mitokondrilerin atasının hakiki ökaryotik hücreler tarafından edinildiği ve ardından mitokondrisi olmayan tüm ökaryotik soyların tükendiği şeklindeki alternatif senaryoyu eleyemeyiz.) Ayrıca, hidrogenozomlara (oksidatif fosforilasyonu olmayan ve hidrojen üretimiyle ATP yaratan işlevsel olarak ayrık organeller) dönüşüm da dahil olmak üzere mitokondriler büyük çapta evrimsel çeşitlenme geçirmiştir (Hjort et al. 2010). Hidrogenozomlar, çeşitli silli protistler ve anoksik deniz çöküntülerindeki lorifiseran hayvanlar da dahil olmak üzere ikincil anaerobik ökaryotlarda birden çok kez evrilmiştir (van der Giezen et al. 2005; Danovaro et al. 2010).
Kloroplastların siyanobakteriyel atası ile ortakyaşam yoluyla oksijenik fotosentezin ökaryotik ediniminin evrimsel tarihi de karmaşıktır. Sadece bir kez evrilen mitokondrilerden farklı olarak, kloroplastlar iki farklı siyanobakteriyel gruptan evrilmiştir (Keeling 2013). Siyanobakteriden-türeme bir kloroplast Synechoccus'un müttefiğidir ve rizaryan (İng. rhizarian) amip Paulinella chromatophora'da raporlanmıştır (Nakayama and Ishida 2009; Nowack and Grossman 2012). Öteki soy ise hem algleri (e.g., Rhodophytes, Chlorophytes) hem de kara bitkilerini kapsayan pek başarılı Arkeplastidlerde bulunur ve bazı alglerin ikincil, hatta üçüncül kloroplast evrimleri olmuştur (Keeling 2013).
Kuşkusuz, ökaryotların evrimsel ve ekolojik başarısının temelinde, ortakyaşamla aerobik solunumu ve fotosentezi edinmeleri ve bakteriyel ortakyaşarların organellere dönüşümü yatar. Bu beceriler olmadan, bir grup olarak ökaryotlar (fotosentetik siyanobakteriler ve aerobik bakterilerle dolu olan) oksik yaşam alanlarına giremezdi; hipoksi ve anoksi tarafından dikte edilen düşük-enerjili bir yaşam tarzıyla sınırlanırdı. Aerobik solunumda ve fotosentezde elektron taşınım zincirinin sırasıyla mitokondriler ve plastidler şeklindeki hücre-içi bölmelere yerelleşmesi, ökaryotlar için ilave evrimsel fırsatlar sağladı. Enerji üretiminin organel zarı ile kısıtlanması, hücre zarının işlevinden ayrılması, büyük hücre boyutuna ve enerji üretimini etkilemeden "hücre zarının yüzey alanı"/"hücre hacmi" oranında azalmaya olanak sağladı (Lane and Martin 2010). Hücre zarına yerelleşmiş solunum zincirli bakteriyel hücreler (bazıları çeşitli uyumlanmalar yoluyla nispeten büyük hücre boyutuna ulaşabilse de) genellikle büyük yüzey alanı:hacim oranıyla küçük boyuta kısıtlanmıştır (Schulz and Jorgensen 2001). Ayrıca, bakteriden-türeme organeller, metabolizmada (örn. yağ asitlerinin β-oksidasyonu, steroid hormon sentezi) ve sinyallemede (örn. apoptoz, kalsiyum sinyalleme) önemli rolleri olan ilave hücre-altı bölmeleri temsil ederek, karmaşık etkileşimleri gerçekleştirmek üzere hücresel verimliliği ve kapasiteyi artırır.
Mikrobiyal Ortakyaşarların Birincil Metabolizması Üzerine Kurulu Ortakyaşamlar
Mikrororganizmalarla ortakyaşam, çağdaş ökaryotların uzak atalarında evrilmiş öbakteriyel içortakyaşarlar (endosimbiyontlar) ile kurulan ortaklıklarla kısıtlanmış değildir. Aslında, bakteriyel ve ökaryotik mikroorganizmaları edinme kapasitesi, ökaryotların biyolojisinde süren bir tema ve ökaryotların farklı yaşam ortamları ile yaşam tarzlarına uyumlanmasının yanı sıra çok sayıda grubun evrimsel çeşitlenmesini biçimlendirmede başlıca unsurdur. Özellikle çarpıcı örnekler, fotosentetik alglerin mantargiller tarafından edinimiyle ortaya çıkan likenlerde ve fotosentetik alglerin hayvan olan mercanlar tarafından edinimiyle ortaya çıkan resiflerde görülebilir; ışığın ulaşabildiği derinliklerde resif oluşturma kapasitesi ortakyaşama bağlıdır.
Ökaryotların ortakyaşam yoluyla elde ettikleri karmaşık metabolik özellikler, aerobik solunum ile fotosentezden ibaret değildir; ama diğer özelliklere katkıda bulunan mikrobiyal ortaklardan hiçbiri organele dönüşmemiştir. Azot bağlama ve kemosentez, atasal ökaryotta bulunmadığı açıkça belli olan iki metabolik özelliktir ve çok sayıda ökaryotik grup tarafından ortakyaşam yoluyla edinilmiştir.
Örosid (İng. eurosid) soyundan kapalı tohumlu bitkiler, başta (Rhizobium ve α-proteobakterilerden Bradyrhizobium dahil) rizobiya (İng. rhizobia) ve Frankia aktinobakterileri olmak üzere azot-bağlayan bakteriler ile ortaklığa özellikle eğilimlidir. Bu bitkilerde atmosferik azota erişim, örneğin ilk ardışık toplulukların azot yoksunu toprakları kullanmalarını sağlar ve azot içeren toksinlerle (alkaloidler dahil) korunan büyük, azot-zengini tohumlarıyla baklagillerin evrimsel çeşitlenmesine katkıda bulunan bir unsur olarak kabul edilir (Houlton et al. 2008; Corby et al. 2011). Azot-bağlayıcı ortakyaşamlar protistlerde, örneğin deniz diatomlarında (Foster et al. 2011) ve bazı hayvanlarda da (odun yiyen gemi kurtlarında ve bazı termitlerde olması dikkat çekicidir) raporlanmıştır (Lechene et al. 2007; Desai and Brune 2012).
Kemosentez (hidrojen, hidrojen sülfit ve metan gibi indirgenmiş maddelerin oksidasyonundan çıkan enerjiyi kullanarak karbon dioksit bağlama kapasitesi), hayvanlar tarafından sıkça kullanılır. Bu ortaklıkların yeşerdiği yaşam alanları, oksik/anoksik karışım bölgeleridir; sucul çökeltiler, hidrotermal bacalar ve hidrokarbon sızıntıları gibi. Bu yaşam alanlarındaki çoğu hayvan, beden yüzeylerinde (örn. kum kurdu), solungaçlarında (örn.çift kabuklu yumuşakçalar) ya da trofozom denilen merkezi dokuda (sakallı solucanlarda) kemosentetik ortakyaşarlar taşır ve çoğu düşük bir beslenme kapasitesi ve indirgenmiş ya da yok olmuş bir sindirim sistemi sergiler (Petersen et al. 2011; Roeselers and Newton 2012).
Çeşitli ökaryotlar, kendilerinin ökaryotik miraslarının metabolik sınırlandırmalarını, daha fazla metabolik beceri yitimiyle birleştirmiştir. Metabolik kapsamdaki azalma özellikle hayvanlarda çok bellidir; bir grup olarak, 20 protein amino asitin (temel amino asitlerin) en az dokuz tanesini ve metabolizma için çok önemli enzimlerin işlevi için gereken çeşitli kofaktörleri (örn. çeşitli vitaminleri) sentezleme kapasiteleri yoktur. Eklembacaklılar, ek olarak, sterol sentezi kapasitesini de yitirmiştir. Genel olarak, bu yitirilmiş metabolik beceriler çok sayıda tepkimeyi içerir ve enerji gerektirir (Wagner 2005). Amino asitler veya kofaktörler içeren bolca besini olan organizmalar, bu becerileri korumakla herhangi bir avantaj kazanmaz (gevşek seçilim) ve belki de onları kaybetmenin enerjik verimliliği seçimsel avantaj sağlar. Bu akıl yürütmelerden öngörüldüğü üzere, süngerler (en temel hayvanlar) ve koanoflajelatlar ( hayvanların en yakın akrabaları) canlı organizmalarla, özellikle de dengeli beslenme sağlayan bakterilerle beslenir.
Hayvanların evriminde çeşitli kereler, mikroorganizmalarla ortakyaşam, hayvanların besinsel olarak dengelenmiş bir diyetle beslenmenin besinsel kısıtlamasından kaçmasına olanak tanımıştır. Hayvanların çok sayıda mikrobiyal ortakyaşarı muhtemelen besinle-ilişkili mikroorganizmaların hayvanlar tarafından sindirimindeki gitgide artan gecikmeyle evrilmiştir; hayvanın besin alımının sindirimden sağlam mikrobiyal hücrelerden salınan biyotrofiğe dönüşümünün eşlik ettiği yetersiz-besinli bir diyete kaydığı evrimsel senaryoya yol açmıştır. Hayvanların mikrobiyal ortakyaşarlardan besinsel desteğe gereksindiği iki diyet, omurgalı kanı ve bitki özsuyudur; yaşam döngüleri boyunca bu diyetlerle beslenen hayvanların tümünün mikrobiyal ortakyaşarlara sahip olması da buna işaret eder (Buchner 1965). Örneğin, zorunlu kan-emicilerden sülük, kene ve tahtakurusunun özelleşmiş hücrelerinde veya sindirim kanalında ortakyaşar mikroorganizmalar doğar ve bunların hayvan konaklarına B vitamini sağladığı düşünülmektedir. Bitki özsuyu ile beslenme de birden fazla kez evrilmiştir; ama sadece Hemiptera takımındaki böcekler (beyaz sinek, yaprak biti, çikada ve yaprak çekirgesi dahil) arasında. Bu böcekler temel amino asitleri, yetersiz miktarda besini bitki soymuk borusu ile odun borusu özsuyundan ve muhtemelen vitaminleri mikrobiyal ortaklarından edinir (Douglas 2009). Ayrıca mikrobiyal ortakyaşam, lignoselüloz dahil olmak üzere yapısal bitki malzemesinden hayvanların yararlanmasında da önemli bir rol oynamıştır. Otçulluk omurgalılarda birden fazla kez evrilmiştir ve mideye alınan selülozdan enerji eldesi, midenin anaerobik bir kısmındaki (fermentasyon odacığındaki) ortakyaşar bakterilerce yapılan selülozize bağlıdır.
Bakteriyel fermantasyonun atık ürünleri kısa-zincirli yağ asitleridir; asetik asit ve bütrik asit gibi. Bu ürünler hayvanın epitel hücrelerinin içine taşınır ve kan yoluyla diğer organlara ulaştırılır; orada aerobik solunum için substrat olarak kullanılırlar (Karasov and Douglas 2013). Mideye alınan lignoselülozun mikrobiyal parçalanması ve selülozun fermentasyonu genellikle omurgasız (özellikle de böcek) otçullar için daha az önemlidir; ama böceklerin (termitler, odun hamamböcekleri, odun arısı ve besinsel içeriği çok düşük olan odun mahsulleriyle beslenen başka bazı böcekler gibi) karbon ekonomisine önemli bir katkı yapabilir (Geib et al. 2008; Suen et al. 2010; Adams et al. 2011). Mikrobiyal ortakyaşarların, yapısal bitki malzemesiyle beslenen böceklere katkılarından daha büyük olan omurgalılara yaptığı katkıya, çok sayıda unsur katkı yapabilir. Bunlar arasında, çoğunlukla anoksik sindirim boşluklu daha büyük bir omurgalıya kıyasla küçük, yüksek düzeyde aerobik bir böcekteki fermentasyon odacığı için daha büyük anatomik engeller olması ve pek çok böcek dahil omurgasızlarda içkin (yani hayvan kökenli) selülazların yaygın varlığına karşın omurgalılarda bunların olmaması da vardır (Davison and Blaxter 2005; Calderon-Cortes et al. 2012). Sonuç itibariyle, omurgalılarda otçulluğun evrimi için mikrobiyal ortakyaşam genel bir ilkedir ama omurgasız hayvanlar için önemi duruma göre değerlendirilmelidir.
Ortakyaşamlar ve İkincil Metabolizma
Ortakyaşar mikroorganizmalar, ökaryotik konaklarının ikincil metabolizmasına da katkıda bulunabilir. "İkincil metabolizma" ifadesi, organizmanın gelişimine ve çoğalmasına doğrudan katkı yapmayan ama organizmanın doğal çevreye uygunluğu açısından kritik önem taşıyabilen metabolik tepkimeleri ve yolakları anlatır. İkincil metabolizma, toksinlerin, antibiyotiklerin, feromonların ve allomonların, pigmentlerin sentezinin yanı sıra, toksinlerin ve diğer ikincil metabolitlerin parçalanmasını da kapsar. Ökaryotların ikincil metabolizma kapasiteleri, yakın akraba türler arasında bile geniş ölçüde çeşitlilik sergiler ve ikincil metabolizma ile çalışmadaki teknik güçlükler, bazı ökaryotların ikincil kimyasında mikroorganizmaların rolünün anlaşılmasında çeşitli başarısızlıkların yanı sıra, bazı kanıtlanmamış ortakyaşarsal ikincil metabolizma iddialarına yol açmıştır.
Birbirini tamamlayan iki yaklaşım, ikincil metabolizmada mikrobiyal ortakyaşarların rolü için harika bir sınama sağlar: Mikrobiyal ortağın dizilenen genomunda ilgili genlerin belirlenmesi ve konaktaki ikincil metabolik kapasitesinin ifadesi ile bu genleri olan mikrobiyal ortağın varlığı arasındaki mükemmel bağlaşıklık (korelasyon). Bu durum, bakteriyel ortakyaşarların, poliketitlerin (biyolojik olarak aktif bir ikincil bileşenler sınıfı) üretimine katkısıyla aydınlatılabilir. Örneğin, Paederus böceği Pseudomonas cinsi bakteriler taşır; bakterilerin bu hayvanların poliketit profilinin üretiminde gereken poliketit sentaz ve bağlantılı tepkimeler için genleri vardır (Piel et al. 2004a) ve Paederus'tan Pseudomonas'ın elimine edilmesi, poliketit üretiminin yitimiyle sonuçlanır (Kellner 2001). Benzer biçimde, pek çok ot, bitki dokuları boyunca dallanan clavicipitaceous fungal endofitler tarafından sentezlenen alkaloidlerle otçullardan korunur (Fletcher and Harvey 1981) ve Lolium otundaki fungal endofitin yaptığı önemli bir alkaloid olan lolinin fungal sentezinin genetik temeli saptanmıştır (Spiering et al. 2005). Mikrobiyal ortakyaşarlar tarafından sentezlenen ikincil metabolitlerin ayrıca süngerler (Piel et al. 2004b), yosun hayvancıkları (Sharp et al. 2007) ve gömlekliler (Kwan et al. 2012) dahil olmak üzere bentik deniz hayvanlarında avcılara ve patojenlere (hastalık yapıcılara) karşı korunmaya katkısı da gösterilmiştir.
İkincil kimya çok geniş bir çeşitlilik gösterdiği gibi, ikincil metabolit sentezinde kullanılan çoğunlukla karmaşık biyosentetik yolaklara ökaryotların erişim kazanma yolları da çeşitlilik gösterir. Bu, hayvanlardaki karotenoid pigmentlerin kaynağına ilişkin bir süre önce yapılan çalışma ile aydınlatılabilir. Karotenoidlerin antioksidan özellikleri vardır ve ayrıca pigment olarak kullanılırlar. Hayvanlar genellikle karotenoidleri sentezleme becerisinden yoksundur ve çoğu hayvan yediklerinden karotenoid alır. Bununla birlikte, bazı böceklerin karotenoid profili yediklerinden bağımsızdır. Beyaz sinekler adındaki bir böcek grubunda, karotenoidler ortakyaşarsal kökenlidir; karotenoid biyosentezi genleri, böceğin bakteriyel ortakyaşarı Portiera tarafından kodlanır (Sloan and Moran 2013). İki başka grupta, yaprak biti ve örümcek akarında, karotenoid biyosentezi genleri hayvanın genomunda kodlanır; bu da mantargillerden hayvanlara yatay gen aktarımı sonucudur (Moran and Jarvik 2010; Altincicek et al. 2012; Novakova and Moran 2012).
İkincil metabolizmanın, üzerinde dikkatlice düşünülmesini hak eden bir yönü daha vardır: Yerleşik mikroorganizmaların ikincil metabolitlerin toksinsizleştirmesine (ağısızlaştırmasına) katkısı. Sindirim mikrobiyotası etkinliklerinin ağız yoluyla alınan ilaçların metabolik yazgısını ve yarı-ömrünü belirleyebileceğine ilişkin kanıtlar ışığında ve ilaç verimliliği ve toksikliği konusunda klinik açıdan önemli sonuçlarıyla, biyomedikal araştırmalarda şu anda bu bir "sıcak gündem maddesi"dir (Haiser and Turnbaugh 2012; Holmes et al. 2012; Haiser et al. 2013). Benzer bir mesele, otçulluk disiplininde onlarca yıldır süren bir ilgi alanı olmuştur. Her ne kadar otçul hayvanlar genellikle çok çeşitli ağısızlaştırıcı enzime (sitokrom P450 monoooksijenazlar, glutation S-transferazlar ve esterazlar da dahil (Despres et al. 2007)) sahip olsa da, bitki allelokimyasallarının ağısızlaştırılmasını gerçekleştimek için uzun süredir ortakyaşar mikroorganizmalara başvurulur. (Jones 1984; Berenbaum 1988; Dillon and Dillon 2004). Birkaç sistemden destekleyici kanıt gelmiştir. Yöreye özgü Endonezya sığırının işkembe mikrobiyotasının, tropikal baklagil Leucaena leucocephala'daki toksik amino asit mimozini parçalama kapasitesinin raporlanmasının gösterdiği üzere, geviş getiren hayvanların önmide fermentasyon odacığındaki (işkembedeki) mikrobiyota, alınan besindeki allelokimyasalları ağısızlaştırma poyansiyeline sahiptir (Cheeke 1994). Benzer biçimde, ren geyiğinin ren geyiği yosununu (bir liken olan Cladonia rangiferina) kullanma yeteneği, ren geyiğinin işkembesindeki bir bakteri olan Eubacterium rangiferina'ya atfedilir; bu bakteri, likende bolca bulunan ve çoğu hayvan için zehirli olan bir metabolit olan usnik asiti metabolize edebilir (Sundset et al. 2008, 2010). Bitki allelokimyasalı ağısızlaştırma, başta ağaç kabuğuyla beslenen böcekler ve yaprak kesen karıncalar olmak üzere bazı böcek ortakyaşamlarında da sezilmiştir. Dağ çamı böceği Dendroctonus ponderosae'nin sindirim mikrobiyotası, terpenlerin (bu böceklerce işgal edilen ağaçların bir numaralı savunma bileşiğinin) parçalanmasıyla ilgili genler taşımakta olan Pseudomonas, Serratia, Rahnella ve Burkholderia cinslerinden bakteriler içerir (Adams et al. 2013). Yaprak kesen karınca Acromyrmex echinatior, yuvasında ortakyaşar mantargil Leucocoprinus gongylophorus yetiştirir ve onunla beslenir. Mantargilin genomu, lakkazlar (bir çeşit fenoloksidaz) için çok sayıda gene sahiptir. Enzimatik açıdan aktif olan lakkaz enzimi, mantargille beslenen karıncaların sindirim kanalından geçer ve bitki malzemesi üstüne salınır; burada, taninler ve flavonoidler gibi bitki bileşenlerini parçalayabilir (De Fine Licht et al. 2013).
Sağlığın Ortakyaşamsal Temeli
Tüm canlı organizmaların işlevi, tek tek hücrelerin içindeki süreçleri koordine eden ve çok-hücreli canlılar için ayrıca hücreler, dokular, organlar arasındaki süreçleri de koordine eden sinyalleme ağları tarafından yönetilir. Bu düzenleyici ağların patojenler (hastalık yapıcılar) ve parazitler (asalaklar) tarafından manipüle edilebileceği, hücre iskeletinin ve hücre-içi trafiğin yeniden düzenlenmesinden, konağın gelişim şablonlarının, üreme zamanlamasının ve davranışının yeniden yapılandırılmasına kadar değişen çeşitli sonuçlarının olabildiği uzun süredir biliniyordu (Gandon et al. 2002; Bhavsar et al. 2007; Hughes et al. 2012). Hastalık yapmayan yerleşik mikroorganizmaların da, ökaryotik konaklarının sinyalleme ağlarını modüle edebildiği ve bu manipülasyonların genellikle konağa avantaj sağladığı giderek daha açık hâle geliyor. Her ne kadar veriler hâlâ parçalar hâlinde olsa da, çeşitli kanıt dizileri, ökaryotların ortakyaşamdan sağlık açısından yarar sağladıkları, çünkü düzenleyici ağlarının, yerleşik mikrobiyota ile etkileşimler bağlamında işlemek üzere yapılandığı hipotezi ile uyum gösteriyor.
Ortakyaşamın sağlık açısından yararları, Azospirillum, Bacillus subtilis, Pseudomonas putida ve Enterobacter cloacae suşlarını da kapsayan bitki gelişimini artıran rizobakteriler (İng. plant-growth-promoting rhizobacteria - PGPR) ile aydınlatılabilir. Adından anlaşılacağı üzere, bu bakteriler (PGPR) bitki kökleri ve bitki gelişiminin artması ile ilişkilendirilir. Altta yatan süreçler karmaşık ve değişkendir, ama çok yaygın olarak sinyalleme molekülleri (bitki hormonları, örneğin auxin indole-3-asetik asit (IAA) ve uçucular, örneğin 2,3-butanediol dahil) sentezleme kapasitesiyle ilgilidir (Spaepen et al. 2007; Lugtenberg and Kamilova 2009; Roca et al. 2013). En iyi incelenmiş olan etkileşim, artırılmış kök dallanmasını ve kök kılların yoğunluk yükselmesini tetikleyen "PGPR'den türeyen IAA" (İng. PGPR-derived IAA) ile ilgilidir. Bu değişmiş kök morfolojisi, topraktan besin emilimini artırır ve böylece bitki büyümesini kolaylaştırır (Dobbelaere et al. 1999; Steenhoudt and Vanderleyden 2000). Bu sonuçların önemli olası sonucu şudur: Bitki gelişim şablonunu kontrol eden düzenleyici ağlar, mikropsuz bitkilerin optimal gelişimini sağlamak üzere yapılanmamıştır; optimal gelişim ancak ilişkili rizosfer bakterilerle sinyal değiş-tokuşu bağlamında başarılır.
IAA üreten PGPR'ler (İng. IAA-producing PGPR), konağın sinyalleme ağının da ayrılmaz bir parçası olan moleküller (bu örnekte, bir bitki hormonu) salarak konak gelişimini etkileyen yerleşik mikroorganizmalara bir örnektir. Mikrobiyal ortakyaşarlar tarafından üretilen diğer bileşikler, konağın sinyalleme ağının bilinen öğeleri değildir ama yine de konak sinyallemesini modüle eder (kuvvetlendirir veya sönümlendirir). Meyve sineği Drosophila ile onun sindirim sistemi mikrobiyotası arasındaki ilişki hakkında yapılan araştırmada bunun bir örneği bulunmuştur. Mikroorganizmalar elimine edildiğinde, hasarlı insülin sinyallemeleri olan sineklerin fenotipik özelliklerine benzer şekilde, Drosophila düşük gelişim hızları ve yüksek yağ düzeyleri ile glukoz dolaşımı sergiler (Shin et al. 2011).
Baskın bir bakteriyel ortakyaşar olan Acetobacter pomorum'un "pirrolokinolin kinona bağlı alkol dehidrojenaz" (İng. pyrroloquinoline quinone-dependent alcohol dehydrogenase - PQQ-ADH) enzimini üretemeyen bir mutantını taşıyan sinekler de bu özellikleri sergiler; insülin-benzeri peptitler (dilp-3 and dilp-5) için olan genlerin ifadesinde de azalma görülür. PQQ-ADH enzimi, etanolün asetik asite oksidasyonunu sağlar. Asetik asit yiyeceklerin içinde olduğunda, mutant bakteriler taşıyan sineklerdeki gelişim hızları, lipid ve glukoz içerikleri ve dilp geni ifadesi, normal sineklerdeki değerlere kayar. Bu veriler, yabani tip A. pomorum tarafından üretilen asetik asitin, sinekte insülin sinyallemesi uyarımı yaptığına işaret eder. Asetik asitin insülin sinyallemesi ile etkileştiği süreçler bilinmemesine karşın, çürüyen meyve üzerinde gelişen ve meyve kaynağı tükenmeden larva gelişimini tamamlaması gereken bu canlının, larva gelişiminde ortaya çıkan hızlanma onun için avantajlıdır. Bitki kökleriyle ilişkili olan "IAA üreten PGPR"lerde olduğu gibi, meyve sineğinin midesindeki "asetik asit üreten bakteri"ler de, konak sinyallemesini yükselterek, konağın sağlığını ve dinçliğini artırır.
Mutant A. pomorum taşıyan Drosophila'da görülen hiperlipidemi ve hiperglisemi, sindirim mikrobiyotasının bileşimini değiştiren antibiyotikler verilen laboratuvar faresindeki yükselmiş lipid ve glukoz düzeylerini anımsatır (Cho et al. 2012). Farenin sindirim mikrobiyotası ayrıca beslenme veya (özellikle besin dağıtımında ve bağışıklık işlevinde olmak üzere, ilgili fenotipik başkalaşımlarla birlikte, özellikle fare bağışıklık sistemi ve besin sinyallemedeki) mutasyonla da değiştirilebilir (Maslowski and Mackay 2011; Claesson et al. 2012; Maynard et al. 2012). Nakil çalışmaları, değişmiş mikrobiyal topluluğun muhtemelen değişmiş konak fenotipine katkıda bulunduğuna işaret ediyor. Mikropsuz (İng. germ-free) kontrol farelerine (standart diyetle yetiştirilen/ yabani tip genotipte) verildiğinde, alıcılar benzer istenmeyen fenotipik özellikler sergiliyor (Vijay-Kumar et al. 2010; Smith et al. 2013). Bu farklı veri kümeleri, yerleşik mikrobiyota bileşiminin ve etkinliklerinin, konak sağlığı için önemli olduğunun kabulünü güçlendiriyor; öyle ki mikrobiyota dengesinin bozulması, "disbiyoz" olarak bilinen kronik sağlık bozukluğuna katkı yapabiliyor. İlk olarak bir yüzyıl önce kullanılan (Metchnikoff 1910) disbiyoz kavramı, muhtemel etmenler olarak çocukların çevresel mikroorganizmalara (hijyen hipotezi) veya yerleşik insan mikrobiyotasının belirli üyelerine (kaybolan mikrobiyota hipotezi) daha az maruz kalmasıyla birlikte, insanlarda bağışıklıksal (immünolojik) ve metabolik hastalıkların artmasını açıklamak için ileri sürülen hipotezler bağlamında anlam kazandı (Strachan 1989; Blaser and Falkow 2009). Disbiyozun uygunluk (İng. fitness) masrafları düşünülürse, hayvanların ve bitkilerin düzenleyici devrelerinin, yerleşik mikrobiyotaları tarafından modülasyona duyarlılığı gayet gerçek bir zafiyet gibi görünür. Nasıl olur da ökaryot-sinyalleme ağları, yerleşik mikroorganizmaların etkisinden yalıtılmayıp, konak-mikrop ortaklaşa kontrolünde gibi görünür?
İki tane süreç kümesi, sağlığın ortakyaşarsal temeline katkı yapabilir. İlki, mikroorganizmaların o anki ve gelecekteki çevresel koşullar için güvenilir bir ipucu sağlamasıdır. Mikrobiyal ürünlere uygun biçimde yanıt veren ökaryotlar, yani büyümede veya gelişimsel şablonlarda, davranışta vb. düzenlenmiş değişiklikler gösterenler seçimsel avantajlı olur. Örneğin, çürüyen meyveyle ilişkilendirilen bakterilerden çıkan metaboli son-ürünlerin profili, meyve kaynağının yaşam süresi için güvenilir bir ipucu sağlayarak, Drosophila larvalarının gelişimsel zamanlamalarını çevresel koşullara göre ayarlamalarına olanak tanır. Özellikle belirtmek gerekirse, Acetobacter'den türeyen asetik asite yanıt olarak yükselen insülin üretimi, gelişimsel hızları artırır ve bu, doğal çevreye yüksek düzeyde uyumlandırıcı olabilir. Mikrobiyal ürünler, çeşitli ökaryotlar tarafından ipucu olarak kullanılır. Örneğin, Algoriphagus machipongonensis bakterisinden salınan bir sulfonolipid, koanoflagellatlarda koloni oluşumunu tetikler (Alegado et al. 2012); Vibrio bakterilerinden salınan N-asil homoserin-laktonlar (yetersayı-algılayıcı moleküller), yeşil alg Enteromorpha'nın hareketli zoosporlarının durgunlaşmasını artırır (Joint et al. 2002); deniz ortamlarındaki substratla ilişkilendirilen Pseudoalteromonas bakterisinden salınan bileşikler, kaya midyesi larvalarının durgunlaşmasını ve başkalaşımını artırır (Hadfield 2011).
Bu değişik bileşenleri ipuçları olarak tanımladım çünkü onların mikroorganizmalar tarafından üretimi, ökaryotik konağın yanıtından bağımsızdır; yani bir ökaryotun mikrobiyal bileşenlerden bilgi elde edip etmediğinin veya nasıl elde ettiğinin, mikroorganizmaların onları üretmesi üzerinde bir etkisi yoktur. Eğer mikroptan türemiş bir bileşen güvenilir bir ipucu ise ökaryotun sinyalleme ağı, diğer çevresel faktörlere karşı azalan yanıt verme yoluyla, bu ipucuna artan bir bağımlılık geliştirebilir ve bu da sinyalleme ağının işlevinin sürdürülmesi için, en sonunda da konak sağlığı için belirli mikrobiyal ürünlere bağımlılıkla sonuçlanabilir.
Konak-sinyalleme ağları üzerindeki mikrobiyal etkilerin alternatif temeli, mikroorganizmalar ile konak arasındaki çekişmeden doğar. Bir ökaryotik konak ile onun yerleşik mikrobiyotası arasındaki eksik seçimsel örtüşmeden dolayı, çekişme kaçınılmazdır. Bir ökaryot, içindeki mikroorganizmaların bağışıklık sistemine tahammül ettiği ve başka savunmaların çoğalabileceği besin zengini bir arazidir ve ayrıca sıklıkla dağılım için bir güzergâhtır. Sonuç olarak, çoğu mikroorganizmanın, konaklarının üreme faaliyetine hiçbir seçimsel ilgisi olmaz. (İstisnai olarak, anneden kalıtılan çeşitli biçimler, konak cinsiyet oranında konakla çekişmededir.) Dolayısıyla, hayvanın sindirim kanalındaki mikroorganizmalar, konak sıhhati için olumsuz sonuçlarına rağmen hiperfajiyi (aşırı oburluğu) destekleyebilir, yerleşiklik sürelerini optimize etmek için konağın optimal besin emilimi pahasına sindirim peristalsis (mide ve bağırsakların sindirim sırasındaki dalgalı hareketi) hızlarını modüle edebilir ve kendi devamlılıklarını sağlamak için bağışıklık yanıtlarını sönümlendirebilir (de La Serre et al. 2010; Round et al. 2011; Matsumoto et al. 2012; Maynard et al. 2012).
Sanılanın aksine, konak-sinyalleme yolaklarının genişliği konusunda konak-mikrop çekişmesi, konak bağımlılığına yol açabilir, ama bu ancak konak popülasyonundaki etkileşen mikroorganizmaların egemenliği çok yüksekse olur. Bu önemli etki, ilk olarak, bir parazitik arı olan Asobara tabida ile dikey iletilmiş bakteri Wolbachia arasındaki ilişki konusunda yapılan bir araştırmada gösterilmiştir. Çeşitli kanıt kümeleri, muhtemelen demir metabolizmasının bozulmasının bir sonucu olarak, Wolbachia'nın oksidatif stres ve apoptotik sinyallemede bağlantılı bir sönümlenme tetiklediğine işaret etmektedir (Kremer et al. 2009). Olası bir sonuç olarak, antibiyotik tedavisi ile Wolbachia'nın elimine edilmesi, yumurtalıkların muazzam apoptozu (programlanmış hücre ölümü) ile sonuçlanarak, arıyı üreme açısından verimsizleştirir. Wolbachia'nın engelleyici manipülasyonlarını kompanse etmek için evrimsel süreçte arının apoptotik yolaklarının yükseltilmiş yanıtlama yeteneği kazandığı iddia edilmiştir (Pannebakker et al. 2007). En önemlisi, bu etki belirleyicidir (tahminen Wolbachia doğal koşullar altında daima varolduğundan) ve uygun apoptotik sinyallemenin Wolbachia bakterilerinin manipülatif müdahalesine gereksinim duyması sonucunu doğurur. Mikrobiyal manipülasyon için benzer konak telafileri, yerleşik mikrobiyota ile hayvanların gelişimsel, besinsel, nörolojik ve immünolojik sağlığı arasındaki etkileşimlerin farklı sergilenişlerine katkı yapabilir (Stappenbeck et al. 2002; Smith et al. 2007; Bravo et al. 2011; Olszak et al. 2012). Ek olarak, sağlığın sürdürülmesi için mikrobiyal ürünlere konak bağımlılığı, çekişmenin sırf sinyalleme ağlarının karmaşıklığı nedeniyle çözüldüğü ökaryotik soylarda korunmuş olabilir. Bir yolaktaki mikrobiyal modülasyonun elimine edilmesi seçilmez, çünkü ilgili sinyalleme yolakları üzerindeki çok sayıda ikincil etki yüksek düzeyde zararlıdır. Sonuç olarak, konak sağlığının ortakyaşarsal temeli, ancak, ökaryotlar ile onların yerleşik mikrobiyotası arasındaki etkileşimlerin derin evrimsel tarihi bağlamında açıklanabilir.
Sonuç Yorumları
Ortakyaşar mikroorganizmalar ile ökaryotların ortaklıklarının öneminin biyologlar tarafından takdir edilmesi uzun süre aldı. Likenlerin (mantargiller ve algler/ siyanobakteriler) ikili doğasının ve bitki kökleri ile mikorrizal mantargiller arasında neredeyse her yerde görülen ortaklıkların 1800'lerde keşfine rağmen, son yüzyılın büyük bir bölümü boyunca, ortakyaşamlar doğanın ilginçliklerinden ibaret görüldü. Mitokondrilerin ve plastidlerin bakteriyel kökenlerine ilişkin artan moleküler kanıtlar, yerleşik mikrobiyotanın insan sağlığı ve ekinler ile büyükbaşların çoğalması için yaşamsal önemde olduğunun anlaşılması, sonunda ortakyaşamı biyolojinin merkezine yerleştirdi. Ama yine de ortakyaşamların evriminin ve işlevinin kavranması için önem taşıyan çok sayıda soru yanıtlanmayı bekliyor.
Yeni dizileme teknolojileri, muazzam fırsatlar sağlıyor. Şu anda büyük ölçüde bakteriyel toplulukların bileşimini ve etkinliklerini kataloglamak için kullanılsa da (Human Microbiome Project 2012b), bu yöntemler hem ökaryotik mikrobiyal ortakyaşarların çeşitliliğini, hem de mikrobiyal toplulukların nasıl yapılandığıını incelemek için uygulanmaya başlıyor (Costello et al. 2012). Önemli sorulardan biri, yerleşik mikroorganizmaların işlevsel özelliklerinin, taksonomik bileşimlerinden öngörülüp görülemeyeceğinin yanıtlanmasını içeriyor. Belki de böyle ilişkiler, mikrop-mikrop ve mikrop-konak etkileşimlerinde sıklıkla karıştırılarak, mikroorganizmaların özelliklerinde mekânsal ve zamansal varyasyonlarla sonuçlanıyordur. Tek-hücre görüntüleme ve tek-hücre dizilemedeki son gelişmeler (Pamp et al. 2012; Pernice et al. 2012; Lasken 2013) böyle heterojenlikleri incelemek için emsali görülmemiş fırsatlar sunuyor. Bu ekolojik meseleler, uzun süredir varolan evrimsel sorulara yavaşça nüfuz ediyor; özellikle belirtmek gerekirse, mikrobiyotanın türleşme şablonlarını ve konaklarının evrimsel çeşitlenmesini etkileyip etkileyemeyeceği sorusu soruluyor (Brucker and Bordenstein 2013). Belirli anlarda, "ortakyaşam adaları" (patojenik bakterilerdeki patojenlik adalarının eşdeğeri) (Finan 2002) yoluyla ortakyaşamla ilgili gen kümelerinin mikroplar-arası aktarımıyla veya bir mikrobiyal ortağın farklı özellikli başka bir taksonla yer değiştirmesiyle evrimsel değişim tesis edilebilir (Koga et al. 2013; Toju et al. 2013). Dramatik bir biçimde ilerleyen genomik teknolojileriyle birlikte, bu temel soruları yanıtlamak giderek daha olası hâle geliyor.
Şu anki araştırmaların çoğunun, ortakyaşamların çeşitliliğinin ufacık bir altkümesine, hayvanlarla özellikle de insanlarla ve laboratuvar fareleriyle ilişkilendirilen birkaç öbakteri şubesine odaklandığının da farkında olmalıyız. Öne çıkan başlıca meselelerden biri, ökaryotların evrimsel yayılımı boyunca ortakyaşamların taksonomik ve işlevsel çeşitliliğidir. Özellikle belirtmek gerekirse, ökaryotların evrimsel çeşitliliğinin büyük bölümünü protistler oluşturmasına rağmen, ister konak ister ortakyaşar olarak olsun, protistlerin dahil olduğu ortakyaşamlar hakkında nispeten az şey bilinmektedir. Bu ortaklıkların bazılarının hayvanlarda ya da bitkilerde paraleli olmadığı açıktır; örneğin iri protistlerin hareket kabiliyeti spiroket dış-ortakyaşarlardan gelir (Cleveland and Grimstone 1964), avcılardan korunma sağlayan dışa çıkık yapılar ise bakteriyel ortakyaşar silli Euplodinium ile sağlanır (Petroni et al. 2000). Protistlerin dahil olduğu ortakyaşamlar hakkındaki artan araştırmalar, ortakyaşamın genel ilkeleri kavramımızı genişletme ve değiştirme potansiyeline sahiptir.- Angela E. Douglas, Symbiosis as a General Principle in Eukaryotic Evolution. Cold Spring Harbor Laboratory Press. doi: 10.1101/cshperspect.a016113
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek OlBunlar da İlginizi Çekebilir





En Çok Okunan
Bu Ay Öne Çıkanlar



İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.