
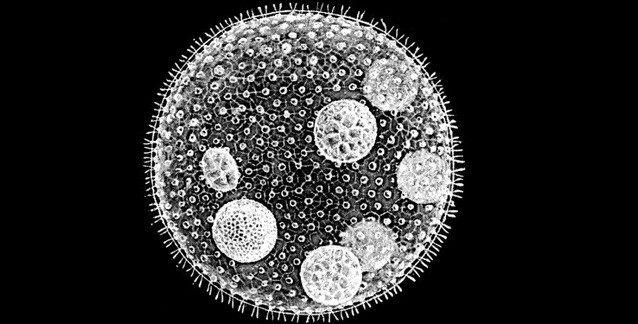

Canlılık, kendini zaman içinde yeniden icat etmekte çok başarılıdır ve en önemli icatlarından biri de çok-hücreliliktir: Yani özelleştirilmiş işlevleri yerine getiren çok sayıda hücre tipi ve hücre yapma becerisi. Çok-hücrelilik evrilmeseydi, gezegenimiz çok farklı bir yer olurdu; bitkisiz ve hayvansız ve elbette insan olmayan bir dünya. Ökaryotik Organizma: Bitki ve hayvanların yanı sıra yeşil algleri, kahverengi algleri, kırmızı algleri, kamçılıları, cıvık mantarları ve mantargilleri de kapsayan. başlıca soylarının çoğunda çok-hücrelilik bağımsız olarak evrilmiş olmasına rağmen, bu evrilişlerin nasıl gerçekleştiği konusunda şaşırtıcı ölçüde az bilgiye sahibiz.
Çok-hücreliliğe sıçrama yapabilmesi için tek-hücreli bir soyun hâlihazırda taşıması gereken belirli özellikler var mı? Böyle bir geçişin gerçekleşebilmesinde özellikle önemli olan belirli çeşitte genler veya gen aileleri ya da genetik mekanizmalar var mı? Çok-hücreliliğin evrimi, genom boyutunda dikkate değer artış ve/veya gen ailelerinde genişleme ya da hatta çok sayıda yeni tür gen gibi büyük adımlar mı gerektiriyor? Yoksa çok-hücreli bir biçime geçiş, sadece ufak değişimlerin gerçekleştiği muhtemelen daha küçük adımlarla mı oluyor? Tek-hücreli Klamidomonas (Chlamydomonas) ile çok-hücreli Volvox'u da içeren bir yeşil alg ailesini inceleyen bilimciler, bu sorulardan bazılarının yanıtlarını bulmaya başladı.
Çok-Hücrelilik Nedir?
Bu sorulara dalmadan önce, çok-hücreli yaşam tarzlarının hepsinin aynı olmadığına dikkat etmek gerekir. Çok-hücreli organizma türlerinin çoğu, gelişimsel mekanizmalar ve geliştirdikleri özellikler açısından birbirlerinden çok farklıdır. Örneğin, tanım gereği çok-hücreli her organizma, hücre bölünmesinin ardından bağlantıda kalan çok sayıda hücreye sahiptir. Ancak bitki ve hayvan türleri, dokular ve organlar şeklinde organize olmuş hücre gruplarını oluşturan en azından bir düzine farklı çeşit hücre üretirken, cıvık mantarlar ve en az bir kamçılı türü gibi bazı çok-hücreli organizmaların çok az sayıda hücre çeşidi olur ve farklı tür hücrelerden oluşan doku ya da organ üretmezler. Dahası, hayvan embriyoları Gastrulasyon: Hücre gruplarının birbirlerine göre konum değişimi yaparken, bitki embriyoları bunu yapmaz. Anlaşıldığı kadarıyla çok-hücreli olmanın çok sayıda farklı yolu var.
Çok-Hücrelilik İçin İyi Model Sistemler Enderdir
Çok-hücreliliğin nasıl doğduğu hakkında bu kadar az bilgimizin olmasının basit bir nedeni var: Araştırılması çok güç olan bir görüngüdür. İşi karmaşıklaştıran etkenlerden biri, bitki ve hayvanlara yol açan geçiş gibi çok-hücreliliğe geçişlerden çoğunun, bir milyar yıl öncesi gibi çok eski tarihlerde gerçekleşmiş olmasıdır. Çok-hücreliliğin ne kadar önce evrildiği neden mesele oluyor? Çünkü var olan tek-hücreli ve çok-hücreli türlerin ortak tek-hücreli atalarından dallanmaları ne kadar eskiden olduysa, çok-hücrelilik ile ilgisi olmayan genetik değişimler de o kadar çok birikmiş oluyor. Biriken bu değişimler, incelenen değişimlerden hangilerinin çok-hücreliliğe geçişte rol oynadığının belirlenmesini güçleştiriyor.
Konuyu zorlaştıran bir diğer etken ise var olan organizmalar arasında karşılaştırma yapmak için iyi birer küme bulmak; yani iyi bir model sistem. Çok az sayıda "tek-hücreli tür & çok-hücreli tür seti" bu çeşit bir kıyaslama yapmaya ve deneysel sınama gerçekleştirmeye uygun, çünkü türlerin birbirleriyle yakın akraba olması gerekiyor. En iyi bilinen hayvan model sistemlerinin (meyve sineği, nematod, zebra balığı, fare vb.) ve bitki model sistemlerinin (Arabidopsis, mısır, pirinç, tütün vb.) bu işe uygun olmamalarının nedeni, onların en yakın tek-hücreli akrabalarının (hayvanlarınki choanoflagellate, bitkilerinki charophycean algi) çok-hücreli soydan çok uzun süre önce ayrılmış olmasından ileri geliyor. Dolayısıyla, hayvanlara ve bitkilere olan genetik benzerlikleri çok az.
İdeal Bir Model Sistem: Volvox, Klamidomonas ve Volvosin Yeşil Algleri
Neyse ki, volvosin yeşil algleri ailesi, çok-hücreliliğin evrimini incelemek için dikkate değer ölçüde elverişli. Volvosin algleri arasında, birbirlerine yakın akraba olan ve günümüzde varlıklarını sürdüren hem tek-hücreli hem de çok-hücreli organizmalar var. Bu gruptaki tek-hücreli türün adı Chlamydomonas reinhardtii ve onunla karşılaştırmaya en uygun olan çok-hücreli yakın akrabasının adı da Volvox carteri.
Bu organizmalar neye benziyor? Klamidomonas, nemli ortamda hareket etmek ve duyusal aktarım için kullandığı iki tepe kamçısı olan tek bir hücreden ibaret. Ancak Klamidomonas hücresinde bu kamçılar her zaman olmuyor. Hücre bölünmesine hazırlanırken onları içine emiyor. Dolayısıyla Klamidomonas yaşam döngüsü, hücrelerin büyüdüğü bir yüzüş evresi ile hücrelerin DNA'larını çoğaltıp bölündükleri bir kamçısız/hareketsiz üreme evresi arasında gidip geliyor. Klamidomonas hücreleri bölünürken, çoklu ayrışma modu denen bir bölünme çeşidi gerçekleştiriyor. Genellikle birbirini izleyen DNA kopyalama ve mitoz turları yaparak, dört, sekiz ve onaltı tane tek-hücreli, cinsiyetsiz yavru hücre üretiyorlar.
Volvox'ta bu iki işlev (yüzme ve üreme), farklı hücre çeşitlerinin görevi oluyor. Volvox'ta somatik hücre denilen çeşitteki hücrelerden yaklaşık 2.000 tane oluyor ve Klamidomonas hücrelerine çok benziyorlar. Bu somatik hücreler küçük ve iki tane kamçıları var; jelatinimsi hücre-dışı matris (İng. extracellular matrix - ECM) küresinin yüzeyinde tek katman hâlinde bulunuyorlar. Bunların görevi, yüzmek ve fotosentez yapabilmesi için Volvox'u ışıkta tutmaktır. Klamidomonas hücrelerinden farklı olarak, Volvox somatik hücreleri bölünemez ve bu ayrım çok önemlidir. Volvox'ta iş bölümü yapılmış bir çok-hücrelilik vardır; çünkü somatik hücreler, üreme becerilerini yitirmiştir. Üreme, gonidium adı verilen başka bir özelleşmiş hücre çeşiti tarafından gerçekleştirilir. Gonidium hücreleri büyük olup, kamçısızdır; dolayısıyla yüzemezler (ve bu yüzden hareket etmek için somatik hücrelere güvenmek zorundadırlar) ama bölünebilirler. Yaklaşık onaltı tane olan gonidium hücrelerinin her biri, yeni bir birey üretme yetisine sahiptir. Bunu, bir sonraki nesilde bulunması gereken tüm hücreleri üreten on ilâ onbir embriyonik hücre bölünmesi dizisi sonucunda yapar. Böyle bir iş bölümü Volvox'a mı özgüdür? Muhtemelen hayır. Bazı bilimciler, somatik işlevler (yüzme gibi) ile üremenin farklı hücre çeşitlerine ayrıştırılmasının, hayvanlardaki çok-hücreliliğin evriminde de yaşanmış olan önemli ilk adımlardan biri olduğunu düşünüyorlar.
Araştırmacılar, günümüzde yaşayan volvosin alglerinin son ortak atasının, modern çağ Klamidomonas hücresine çok benzeyen bir tek-hücreli olduğunu düşünüyorlar ve Klamidomonas'ın genetik düzeyde bu atadan çok fazla farklılaşmamış olabileceğini düşünüyorlar. Araştırmacılar ayrıca bitki ve alg fosillerinden elde edilen bilgilere ve moleküler saat analizlerine dayanarak, volvosin ailesinin üyelerinin birbirlerinden sadece 200 milyon yıldır farklılaştıklarını biliyorlar. Hem Volvox hem de Klamidomonas'ın hem genetik hem de moleküler düzeyde manipüle edilebilen iyi deneysel organizmalar olduğu gerçeği de düşünüldüğünde, bu türlerde çok-hücreliliğe olanak tanıyan genetik yenilikleri keşfetmenin uygun olacağı görülebilir.
Volvosin Alglerinde Çok-Hücrelilik
Geçirilen evrimsel süreçler bakımından, Volvox ile bitkiler, hayvanlar ve diğer çok-hücreli organizmaların kıyaslaması nasıldır? Bir açıdan, Volvox nispeten sınırlı tipte bir çok-hücrelilik sergiler. Sadece iki hücre çeşidine sahiptir ve bu hücreler doku ya da organ şeklinde organize değildir. Yine de etkileyici derecede bir gelişimsel ve morfolojik yenilik evrimleştirmiştir. David Kirk tarafından Volvox, Klamidomonas ve diğer birkaç volvosin algin gelişimsel programları karşılaştırılmış ve tek-hücreli atada olmayan oniki yeni gelişimsel özelliğin Volvox'ta evrildiği sonucuna ulaşılmıştır. Örneğin, Volvox embriyosu tersinme sergiler; bu, hayvanlardaki gastrulasyona benzeyen ve hücre bölünmesini takiben somatik hücrelerin kamçı uçlarını konumlandıran bir morfogenetik süreçtir. Ek olarak, Volvox embriyoları, asimetrik bölünme adı verilen ve farklı büyüklük ve çeşitte hücreler üreten özel bir hücre bölünme çeşiti uygular. Yukarıda tanımlandığı gibi, Volvox özelleşmiş hücre çeşitleri yapar: Fâni somatik hücreler ve ölümsüz kök hücre benzeri gonidiumlar. Araştırmacılar, bu özelliklerin her birinin nasıl evrildiği hakkında birkaç ipucu keşfetti bile.
Çok-Hücreliliğin Evrimini İncelemek İçin Stratejiler
Çok-hücreliliğin nasıl evrildiği nasıl araştırılır? Çok-hücreliliğin genetik temelini incelemek için araştırmacıların kullandığı yaklaşımların çoğu, karşılaştırmalı genomik ve mutasyonel/işlevsel genetik adlı iki stratejiden biri ya da ikisinin bir kombinasyonu olur. Üçüncü bir yaklaşım ise türlerin akrabalık ilişkileriyle ilgili olan moleküler taksonomi çalışmalarıdır ve çok önemlidir. Aslında bu üçüncü yaklaşım, diğer iki yaklaşımın üzerine inşa edildiği temeli teşkil eder. Bu yazıda bu yaklaşıma değinmeyip, onun oluşturduğu temel üzerindeki karşılaştırmalı genomik ve mutasyonel/işlevsel genetik yaklaşımlarına bakacağız.
Karşılaştırmalı genomik yaklaşımlarında, araştırmacılar yakın akraba olan tek-hücreli ve çok-hücreli kuzen türlerin eksiksiz olarak dizilenmiş genomlarını kıyaslayarak, her bir genoma özgü olan genleri ve genomlar tarafından kodlanan proteinlerdeki farkları belirler. Bu tür bir çözümlemenin ardında yatan fikir şu ki, çok-hücreli türde bulunup tek-hücreli türde bulunmayan herhangi bir gen ya da gen ailesi, çok-hücreliliğin evriminde önemli rol oynamış olabilir. Yani çok-hücrelilik için olan bazı "özel" genler sadece çok-hücreli organizmalarda olabilir. Ya da eğer çok-hücreli türde belli bir gen çeşidinden dikkate değer ölçüde fazla kopya varsa veya eğer bazı ilişkili genler tarafından kodlanan proteinler iki türde önemli ölçüde değişmişse, o zaman o fazla gen kopyaları veya değişmiş proteinler, çok-hücrelilik için önemli olmuş olabilir.
Büyük-ölçekli kıyaslamalı genomik incelemelerinin tipik olarak sadece gen ailelerindeki büyük farklılıkları veya iyi bilinen gen ya da gen ailelerindeki farklılıkları ortaya çıkardığını akılda tutmak önemlidir. Böyle incelemeler, küçük farklılıkları açığa çıkaramayabilir; örneğin türlerden sadece birinde bir gen ikilendiği (ya da yitirildiği) zaman, gen ailesi büyüklüğünde oluşan küçük değişim gibi. Evrimsel canlıbilimciler, yeni özelliklerin evrimi açısından gen ikilenmesi olaylarının büyük önem taşıdığını düşünüyor, çünkü yeni genler zamanla değişebilir oluyor ve böylelikle kopyalandıkları genlerden daha farklı işlevler kazanabiliyorlar.
Araştırmacılar ayrıca tek-hücreli türlerde olmayan gelişimsel süreçler (farklı tip hücre yapma becerisi gibi) için kötü olan mutant çok-hücreli versiyonların tanımlanması ile başlayan mutasyonel/işlevsel yaklaşımlar da kullanıyor. Bu mutantlar daha sonra etkilenen genleri klonlamak için kullanılıyor. Ardından araştırmacılar tek-hücreli türlerin genomlarını çözümleyerek, aynı (ortolog) genlerin var olup olmadığını belirliyor ve eğer varsa çok-hücreli versiyondan farklı mı, farklıysa nasıl farklı diye bakıyorlar. Şu anda yaşayan canlılar kullanılarak yapılan bu tür incelemeler çok yararlı oluyor. Yeni becerilerin evrimleşmesinde hangi genlerin önemli olduğuna ilişkin değerli ipuçlarını açığa çıkarıyorlar ve bu genlerin evrimsel tarih boyunca değişimlerini anlamamızı sağlıyorlar. Sonuç olarak, çok-hücreli ve tek-hücreli organizmalar arasındaki farklılıkları listelemek, çok-hücreliliğin nasıl evrildiği konusunda kavrayışımızı geliştiriyor.
Volvosin Alglerinden Öğrenilenler: Karşılaştırmalı Genomik
Volvosin algleri, çok-hücreliliğin nasıl evrildiği hakkında bize ne öğretti? Araştırmacılar kısa süre önce Klamidomonas ve Volvox genomlarını diziledi ve kıyasladı. Bu karşılaştırma sonucunda, dikkate değer ölçüde benzer oldukları görüldü. Neredeyse her açıdan (tüm genom büyüklüğü, protein kodlayan gen sayısı, kodlanan farklı türde protein alanı türü sayısı ve gen ailesi büyüklüklerinin dağılımı) bu iki organizma hemen hemen aynıydı.
Araştırmacılar belirli gen ailelerine daha dikkatli baktıklarında, özellikle de Volvox'ta gerçekleşip Klamidomonas'da gerçekleşmeyen gelişimsel süreçlerin düzenlenmesi ile ilgili oldukları bilinenlerin, büyük ölçüde benzerlikler buldular. Bununla birlikte, bir tane açıkça belli ve önemli farklılık buldular: Klamidomonas'a kıyasla Volvox'ta hücre duvarı/ECM proteinlerini kodlayan çok daha fazla gen vardı ve fazlalık genlerin çoğu, Klamidomonas'ın sahip olduklarından oldukça farklıydı. Bu aşamada Klamidomonas'ı çevreleyen hücre duvarının iki kısmı olduğunu vurgulamak önemlidir: İç katman ve dış katman. Volvox'ta her ikisinin de versiyonları bulunuyor ama iç katmanı, Klamidomonas'ın iç katmanına kıyasla fazlasıyla genişlemiş. Bu da Klamidomonas'da bulunmayan ECM yığınını oluşturuyor ve Volvox hücrelerinin birbirlerine yapışmasına yardım ediyor. Araştırmacılar, hücre duvarı genlerindeki patlamanın ve bu genlerden bazılarının farklı tür hücre duvarı genleri hâline gelmelerinin, Volvox'taki ECM'nin ortaya çıkışını tetikleyen şey olduğunu düşünüyorlar.
Volvosin Alglerinden Öğrenilenler: Mutasyonel/İşlevsel Genetik
Açıkça belli ki, saf karşılaştırmalı genomik yaklaşımlarının sınırlandırmaları var; gelişimsel süreçlerin ve çok-hücreliliğin nasıl evrildiği hakkında bilinecek her şeyi bize söyleyemezler. Ama Volvox ve Klamidomonas için genetik taramalar mümkündür. Volvosin soyunda çok-hücreliliğin nasıl evrildiği konusunda bu taramalar hangi bilgileri verir?
Araştırmacılar, Volvox'ta gelişimsel hatalar için genetik görüntüleme kullandı; böylece asimetrik bölünme için gereken bir gen (glsA) ve embriyo tersinmesi için gereken üç başka gen (invA, invB, invC) belirlediler. Bu dört genin hepsinin Klamidomonas'ta kolayca tanınabilen, Volvox'taki karşılıklarına çok benzeyen ortologları vardı. Bilimciler, iki Volvox gelişim genine karşılık gelen Klamidomonas ortologlarını klonladı. Volvox'un glsA geninin ortologu olan Klamidomonas'ın GAR1 geninin, tıpkı glsA gibi işlev gösterebildiği bulundu: glsA mutantlarına dönüştürüldüğü zaman, onların asimetrik bölünme hatasını onarıyor veya kurtarabiliyordu. Benzer biçimde, Volvox'taki invA geninin ortologu olan Klamidomonas'ın IAR1 geninin, invA mutantlarının tersinme hatasını kurtarabildiği bulundu. Bu sonuçlar bize şunu söylüyor: glsA/GAR1 ve invA/IAR1 genleri, Volvox ile Klamidomonas'ın ortak bir atadan dallandığı zamandan bu yana, önemli ölçüde değişmeden kalmıştır.
Peki ama bu genler Volvox gelişiminde böylesine kritik rol oynarken, bu nasıl olabilir? Var olan genlerin (glsA ve invA gibi) değişmeden yeni gelişimsel yolaklara uydurulmasını, bir benzetim üzerinden düşünebiliriz: Benzinli-elektrikli hibrit otomobil. Her arabada fren olur. Hibritler, frenleme sırasında üretilen potansiyel enerjiyi elektriğe dönüştürecek şekilde tasarlanmıştır. Hibritlerdeki frenler hâlâ fren vazifesi yapar ama buna ilaveten, elektrik üreten yeni bir "yolak"a da uydurulmuşlardır. Hibrit arabadan frenleri çıkarırsanız, elektrik üretemez olur. Bu benzetimdeki frenlerin yerinde glsA ve invA genlerinin olduğunu düşünün. Volvox'un tek-hücreli atasındaki işlevlerine hâlâ sahiptirler ama onları çıkarırsanız, Volvox asimetrik bölünme veya tersinme yapamaz olur.
Somatik yenileyici (rejeneratör) regA geninin analizinden de bazı ilave kavrayışlar edinilebilir. Bu gen, Volvox'ta somatik hücrenin yazgısının sürdürülmesi için gereklidir. Mutant regA olan somatik hücreler başlangıçta normal gelişir ama tüm yaşamları boyunca somatik hücre olarak kalıp sonra da ölmek yerine (somatik hücrelerin normalde yaptığı budur), genişleyip gonidiumlar gibi çoğalırlar. O halde, regA geni bir şekilde somatik hücrelerin büyümesini ve bölünmesini engellemektedir ve onların gonidiumların sahip olduğu kök-hücre benzeri potansiyele sahip olmasına izin vermemektedir. Kanser hücrelerinde görülen kontrolsüz büyümeyi önleyen bir tümör baskılayıcı gen olarak regA genini düşünelim. Kaç tane regA benzeri gene sahip olduklarını belirlemek için Volvox ve Klamidomonas'ın genomları çözümlendiğinde, bilimciler her iki algin de, regA ürününü anımsatan proteinler kodlayan büyük bir paralog genler ailesine sahip olduklarını keşfettiler. Ama filogenetik analizler ve başka yöntemler kullanılarak, Klamidomonas'ın bir regA genine sahip olmadığını da buldular. Neden yok? Ayrıca regA geni ilk olarak nereden gelmişti ve somatik hücre yazgısının başlıca düzenleyicisi rolüne nasıl sahip olmuştu?
Klamidomonas ve Volvox genomlarının arkeolojik analizlerini daha da ilerleterek, bilimciler bu sorulardan bazıları için yantlar buldu. Yaptıkları çözümlemeler sonucunda, Klamidomonas ve Volvox'un atasındaki öncül bir gen yanlışlıkla kopyalanarak iki paralog gen ürettiğinde regA'nın ortaya çıkmış olmasının muhtemel olduğu anlaşıldı. Paralog genlerden biri nihayetinde regA olurken, diğeri de akraba bir gen ortaya çıkardı. Volvox hem regA genini, hem de diğer geni (paralogunu) korurken, Klamidomonas ise regA'sını yitirdi. İşlevlerinin nasıl evrildiği konusunda ise ipucu aranacak en iyi yer, diğer genin modern çağ versiyonları olsa gerek. Bu sorunun peşine düşen bilimciler, bu regA benzeri genin Klamidomonas versiyonu olan RLS1'in, Klamidomonas ışıktan ve belirli besinlerden mahrum kaldığında aktifleştiğini buldular. Bu bağlaşıklık, RLS1'in hücreler enerjiden ve besinden mahrum kaldığında işliyor olabileceğine işaret ediyor. Üreme regA tarafından baskılandığında, RLS1'in de öyle yapıyor olması mantıklı geliyor.
Eğer durum buysa, o zaman farklı hücre çeşitleri yapma kapasitesi, enerji/besin mahrumiyetine yanıt olarak hücre büyümesi ve bölünmesini baskılayan bir yolaktan evrilmiş olabilir. Bu olay, bu yolağı kontrol eden gen kopyalanıp, tüm yolağı büyümeyi ve bölünmeyi baskılamak üzere kullanmaya başlayınca gerçekleşmiş olabilir. Hibrit otomobil benzetimini yeniden düşünelim ama bu kez tüm stres yanıt yolağı, fren sistemi olsun. Bunun gibi bir olay (var olan bir genetik yolağın ayarlanmasıyla, hücrenin hep yaptığı bir şeyi farklı koşullar altında yapmaya başlaması) organizmaların nasıl yeni hücre çeşitleri evrimleştirdiğini genel olarak açıklayabilir.- Volvox, Chlamydomonas, and the Evolution of Multicellularity https://www.nature.com/scitable/topicpage/volvox-chlamydomonas-and-the-evolution-of-multicellularity-14433403
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek OlBunlar da İlginizi Çekebilir






En Çok Okunan
Bu Ay Öne Çıkanlar



İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.