
Öz
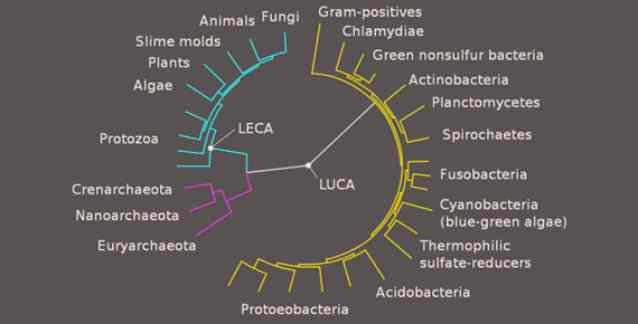
Canlılığın Kökeni Ortamı (CKO; İng. Origin of Life Domain - OLD), Dünya üzerinde canlılığın başladığı dönemdir. Biz burada, CKO’nun evrilmesini sağlayan kronolojik adımları yeniden yapılandırmak amacıyla, Protein Aileleri’ni çözümlemek için yeni bir filogenetik algoritma geliştirip kullanıyoruz. Bu dönem sırasında, RNA’lar, DNA’lar, amino asitler ve zarlar gibi canlılığın temel bileşenlerinin belirmesiyle birlikte canlılık başlamıştır. Canlılığın Kökeni kronolojik olarak Son Evrensel Ortak Ata’dan (SEOA; İng. Last Universal Common Ancestor - LUCA) önce gelir; SEOA daha sonra Dünya üzerindeki modern canlılığa yol açmıştır. Filogenetik algoritmamız, eskiden bilinmeyen canlılığın kökeni sorularını açıkça yanıtlamamıza olanak tanıyor. Özellikle belirtmek gerekir ki, RNA ve DNA dünyalarının evrimini tanımlayan halkaları yeniden yapılandırarak, kullandığımız bilgi-işlemsel yöntemleri açıklıyor ve gösteriyoruz. Filogenetik olarak, RNA ve DNA dünyalarının nasıl evrildiğini yeniden yapılandırıyor, ilk genetik kodların kökenlerini ve belirişlerinin kronolojik sıralamasını çıkarıyor, Ribozomal RNA dünyasının Zar dünyasından önce gelip gelmediğini sınıyor ve bu yeni bulguları başkaları tarafından yapılan deneysel ve kuramsal canlılığın kökeni araştırmalarına göre yorumluyoruz.
Giriş
Canlılığın evrimi giderek daha ayrıntılı biçimde incelenirken, canlılığın ilk zamanlarına ilişkin kavrayışımız da filogenetik yeniden-yapılandırmalar sayesinde giderek daha eski zamanlara çekiliyor (Cox et al. 2008; McInerney et al. 2014). Önceki zamanlarda yapılan filogenetik incelemeler, Dünya’da var olan tüm yaşama yol açan son evrensel ortak ata olan SEOA dönemindeki canlılığın önemli özelliklerini yeniden yapılandırdı (Koonin & Martin 2005; Martin et al. 2008). Ancak SEOA’dan önce gelen Canlılığın Kökeni Ortamı’ndaki (CKO) evrimi açık bir şekilde yeniden yapılandırmanın olanaksız olduğu düşünülüyor.
CKO’yu yeniden yapılandırmanın bu olanaksızlığına rağmen, dolaylı fakat oldukça verimli bir yol izleyerek, canlılığın kökeni araştırmaları yine de ilerledi. Bilimciler, canlılığın ilk zamanlarında var olan yaşamın birincil moleküllerini üretebilecek biyokimyasal tepkimeleri deneysel olarak aradı. Bu çalışmalar, aşağıda tanımlandığı gibi, Dünya’daki canlılığın evriminin ilk zamanlarında büyük olasılıkla var olan molekülleri (nükleik asitler, zarlar, amino asitler ve ribozomlar dahil olmak üzere) üreten çeşitli in vitro biyosentetik yolaklar keşfetmeyi başardı.
Deneysel canlılığın kökeni çalışmaları, Dünya’nın erken döneminde var olduğu düşünülen indirgeyici atmosferik koşullar altında elektriksel kıvılcım boşalımları kullanarak amino asitler sentezleyen Miller-Urey deneyleriyle (Miller 1953) başlamıştı. Bu deneylerin ardından “RNA Dünyası” önerisi geldi (Gilbert 1986). Her iki yaklaşım da nihayetinde araştırmacıları, ilk amino asitleri, nükleik asitleri ve zarları üretmiş olabilecek Dünya’daki ilkel kimyaları keşfetmeye itti.
Son yirmi yılda, canlılığın kökeni tepkimelerinin biyokimyasal arayışları daha önceki sonuçları genişleterek, erken dönemdeki biyolojik tepkimelerin (Mansy & Szostak 2008; Powner et al. 2009) ve SEOA döneminde canlılığın erken evre evrimini büyük olasılıkla etkilemiş olan potansiyel ortamların (Weiss et al. 2016) kapsamlı bir kavrayışını ortaya koydu. Bu başarılara karşın, yaşam bilimcilerinin çoğu, genomik ve dizilimsel veriler kullanarak SEOA öncesindeki canlılık evrimini yeniden yapılandırmanın olanaksız olacağını düşünüyordu. Eğer canlılığın kökenini filogenetik olarak yeniden yapılandırmak olanaklı olsaydı, o zaman biyolojik evrimin bu en erken evresine ilişkin bilgimiz muazzam artardı.
Biz burada Sanger Pfam veritabanını (~12 milyon dizilim, 16.245 Protein Ailesi ve 35 milyon UniProt Tanımlayıcı içerir) kullandık ve zarların, RNA dünyasının ve DNA dünyasının evrildiği evrimsel adımları nasıl açık şekilde yeniden yapılandırdığımızı gösterdik. Genomik filogenetik yeniden-yapılandırma yöntemlerimizi kullanarak, genetik kodun evrimindeki adımları yeniden-yapılandırdık (P < 0,05) ve ribozomal RNA dünyasının RNA/zar dünyasından önce geldiğini ortaya koyduk (P < 0,05).
Canlılığın Kökenindeki Kilometre Taşlarının Yeniden-Yapılandırılması
Son on yılda, SEOA-sonrası dönemde var olan genleri yeniden yapılandırmak için ağaç-bazlı filogenetik yöntemler kullanıldı. Bu süreçte, genom evriminin hem ıraksamalarla (ağaç-benzeri dallanmalarla), hem de yakınsamalarla (gen akışlarının halka oluşturacak şekilde birleşmesiyle) ilerlediğini öğrendik. Biz burada, ilk RNA’ları, DNA’ları ve zarları üreten canlılığın kökeni gen akışlarının kronolojisini filogenetik olarak yeniden yapılandırmak için halka bazlı tekniklerin Pfam verilerini nasıl kullanabileceğini gösteriyoruz.
Fig.1’de gösterilen karmaşık gen akışları, Dünya’daki canlılığın evrimindeki üç evreyi tanımlıyor. En üstte gösterilen halkalar Protein Aileleri’nden yani Pfam’lardan (ortak bir evrimsel kökeni paylaşan protein bölgeleri) yeniden yapılandırıldı. Tekil Pfam’lar, benzer dizilimlerinden, yapılarından ve işlevlerinden tanındı. Protein aileleri karmaşık yapıları temsil ettiği için nükleotitlerden veya amino asitlerden çok daha yavaş evrilirler. Bu da onları, derin (ilk canlılığın kökeni “organizma”larından SEOA’ya ve ondan da Canlılık Halkaları’na (CH; İng. Rings of Life - ROL) yaşamın evrimini yeniden-yapılandırmak için burada kullanılanlar gibi) evrimsel yeniden-yapılandırımlar gerçekleştirmek için ideal yapar. Bu gen akışları yakınsayarak halkalar ve ıraksayarak ağaçlar oluşturabildiğinden, Dünya’daki yaşamın evrimi sırasında gerçekleşen başlıca evrimsel olayların eksiksiz bir matematiksel temsilini verirler (Lake et al. 2015). SEOA, Canlılık Halkaları’ndan önce gelmiş ve onlara yol açmıştır. SEOA’nın moleküler incelemelerinin sonucunda, SEOA’da hangi Pfam’ların bulunduğunu artık oldukça ayrıntılı olarak bilebiliyoruz (Weiss et al. 2016).

Son on yılda, filogenetik yeniden-yapılandırımlar, Dünya’daki yaşamın yakın zamandaki evrimini tanımlayan evrimsel halkalar için kanıt sağladı. Çoğu gen bilinen tüm organizmalarda ortak olduğundan, var olan yaşamın SEOA’dan evrilmiş olduğu genel kabul görmektedir.
Ancak canlılık var olmaya SEOA’dan başlamamıştır ve SEOA’dan önce geniş çeşitlilikte bir grup organizma var olmuştur. Fig.1’deki CKO, canlılığın ilk zamanlarında var olmuş olan bu organizmaları kapsar. CKO, canlılığın ilk belirdiği ve DNA’lar, RNA’lar, zarlar ve amino asitler gibi yeni biyokimyasal icatları kullanmaya başladığı dönem sırasında yaşamın evrimini tanımlar.
Canlılık halkalarını Pfam’lardan yeniden yapılandırmak nispeten kolay olsa da (Lake et al. 2015) ve SEOA’da bulunan protein ailelerinin çoğu bilinse de (Weiss et al. 2016), CKO’daki yaşamın filogenisi hakkında çok daha azı bilinmektedir.
SEOA’nın bizim CKO’yu yeniden yapılandırmamızı engellediği, çünkü Canlılık Halkaları ile CKO arasında durduğu düşünülüyordu; yani onun konumu bizim CKO halkalarını görmemizi önlüyor gibi görünüyordu. Neyse ki, durum böyle değil. İlerleyen bölümlerde, CKO’daki yaşamın yeniden yapılandırılması için SEOA’nın nasıl kullanılabileceğini tanımlıyor, CKO’daki evrimin nasıl yeniden yapılandırılacağını gösteriyor ve CKO’nun bu ilk yeniden-yapılandırımlarını Dünya’daki yaşamın kökenine ilişkin güncel bilgilerimiz ışığında yorumluyoruz.
Malzeme ve Yöntemler
CKO’yu yeniden yapılandırmak için Protein Aileleri’nin evrimini regüle eden evrimsel süreçleri anlamak önemlidir. Özellikle belirtmek gerekir ki, gen akışlarının evrimini doğrudan etkileyen popülasyon bazlı evrimsel mekanizmaları anlamak temel önemdedir çünkü bu mekanizmalar, organizma topluluklarının yeni protein ailelerini kolayca edinmelerine olanak tanır ve aynı zamanda gen akışlarıyla Pfam’ların kaybedilmesini önler (Lake et al. 2015). Fig.2 bunun nasıl olduğunu göstermektedir.

Fig.2’nin solundaki büyük siyah daire, başlangıçtaki gen akışında mevcut olan ayrı Pfam’ları temsil eden üç yeşil + işareti içerir. Zaman içerisinde bu topluluktaki bazı bireyler, bu gen akışındaki bazı Pfam’ları rastlantısal olarak kaybeder (üstteki dikdörtgen içindeki siyah dairelerde eksik olan yeşil artılar). Bu arada, başka bireyler de ortamdan ve başka organizmalardan yeni genler edinir (alttaki dikdörtgen içindeki siyah dairelere eklenen kırmızı artılar). Dolayısıyla, üstteki dikdörtgenin yanındaki büyük daire, gen akışındaki tüm organizmalarda bulunan farklı Pfam’ların toplam sayısını temsil eder. Tekil organizmaların bazıları, hatta çoğu genleri kaybetse bile, o genler yine de dikdörtgenle temsil edilen popülasyonda kalır çünkü büyük bir toplulukta aynı bir genin tüm kopyalarının tüm organizmalar tarafından yitirilmesi matematiksel olarak olanaksız olurdu. Bu, canlılık halkalarında özellikle doğrudur çünkü bazı prokaryotik şubelerin 1.029 ya da daha fazla birey içerdiği tahmin edilmektedir (Whitman et al. 1998).
Bunun tersine, alttaki büyük dikdörtgenle gösterildiği gibi, filogenetik akışların gen kazanması kolaydır. Eğer yeni bir Pfam (Fig.2’de kırmızı bir + ile gösterilmiştir) içeren bir gen topluluğun tek bir üyesi tarafından, örneğin bir Alfaproteobakteri tarafından bile kazanılsa, bu gen Pfam aramalarında Alfaproteobakterilerde “mevcut” olarak tanımlanacaktır. Dolayısıyla zaman içinde, Alfaproteobakterilerde bulunan ayrı Pfam’ların sayısı, bireyler yeni Pfam’lar kazandıkça sürekli artacaktır. O hâlde, yitiren hücrelerdeki Pfam koleksiyonu ile kazanan hücrelerdeki Pfam koleksiyonu toplandığında net sonuç, zaman içindeki gen akışında bulunan Protein Aileleri sayısında süren bir artıştır; bu da Fig.2’de edinilmiş (kırmızı) genlerin artışı ile gösterilmiştir.
Yaptığımız CKO yeniden-yapılandırımları, gen akışları içindeki Pfam’ların sayısının zamanla sürekli arttığına ilişkin bu gözleme dayanmaktadır. Bundan yapılacak çıkarım, daha eski zamanlara bakıldıkça, gen akışındaki Pfam’ların sayısının azalacağıdır. Sonuç olarak, CH gen akışlarında çok sayıda Pfam, SEOA’da daha az Pfam ve CKO (en eski) gen akışlarında çok daha az Pfam olacaktır.
CKO (Canlılığın Kökeni Ortamı), SEOA (Son Evrensel Ortak Ata) ve CH (Canlılık Halkaları) içindeki gen akışlarının toplamı, canlılığın kökeninden son evrensel ortak ataya ve ondan da canlılık halkalarına doğru canlılığın evrimini yeniden yapılandırmak için gereken tüm kronolojik bilgiyi içerir. En eski Pfam gen akışları, en küçük Pfam sayıları ile desteklendiğinden, en küçük gen akışlarını içeren grup olarak istatistiksel olarak tanımlanabilir; tıpkı daha önce en büyük gen akışlarını modern Canlılık Halkaları üyeleri olarak tanımladığımız gibi (Rivera & Lake 2004). Dolayısıyla, en küçük akışlar, CKO içindeki evrimi yeniden yapılandırmak için gereken bilgiyi içerir.
CKO’daki Organizmaların Kategorize Edilmesi ve Etiketlenmesi
CKO’daki evrimi yeniden yapılandırmak amacıyla, ilk olarak CKO’daki organizmaları, var olan organizmaların takımlarına benzer şekilde gruplara ayırıp etiketledik ve kategorize ettik; daha sonra etiketlenen bu grupları analiz etik.
Canlılığın kökeni organizmaları modern yaşamdan milyarlarca yıl eski olduğundan, canlılığın ilk belirmekte olduğu sırada var olduğu genel olarak düşünülen moleküllere göre onları sınıflandırdık. Canlılığın kökeni organizmaları olduğundan en çok kuşkulanılanlardan bazıları, amino asitleri, DNA’ları, RNA’ları ve zarları kullananlardır (Miller 1953; Gilbert 1986; Koonin & Martin 2005; Mansy & Szostak 2008; Martin et al. 2008). Örneğin, eğer bir canlılığın kökeni organizması RNA, Zarlar ve DNA içeriyorsa, onu bir “RNA, DNA, Zar” organizması olarak sınıflandırıyoruz. Benzer biçimde, RNA ve Zarlar kullanan CKO organizmalarını da “RNA, Zarlar” kategorisine yerleştirerek etiketleyip sınıflandırıyoruz. Daha sonra CKO organizmalarını etiketlemek ve Pfam’lardan onların evrimsel yollarını yeniden yapılandırmak için (Canlılık Halkaları’nın Pfam’lardan yeniden yapılandırılmasına benzer şekilde) bu sistemi kullanıyoruz.
Canlılık Kökeni Halkalarının Yeniden Yapılandırılması
Biz burada, Tablo.1’de gösterilen üç tane Pfam varlığı/yokluğu verileri kümesinden, Canlılığın Kökeni halkalarını filogenetik olarak yeniden yapılandırıyoruz. Aşağıda listelenen üç adet Pfam verileri kümesi, Pfam tanımlayıcıların belirtilen varlık/yokluk örüntülerini kaç kez içerdiğinin sayısını temsil etmektedir. İlk veri kümesinin ilk satırı, dört terimin hepsinin (“rRNA,” “mRNA,” “tRNA,” “Zarlar”) 49 farklı Pfam’da görüldüğünü gösteriyor; ikinci satır 15 Pfam’da üç terimin (“rRNA,” “mRNA,” “tRNA”) olduğunu gösteriyor.
Aşağıda verilen üç varlık/yokluk tablosu, üç farklı canlılığın kökeni halkasını yeniden yapılandırmamızı sağlıyor. Tablo.1: Canlılığın Kökeni Halkası Analizleri için Örüntü Sayımları
| rRNA, mRNA, tRNA, Zar / | / Adenin, Guanin, Urasil, Sitozin / | / Adenin, Guanin, Timin, Sitozin |
| + + + + 49 | + + + + 7 | + + + + 4 |
| + + + − 15 | + + + − 5 | + + + − 1 |
| + + − + 7 | + + − + 14 | + + − + 17 |
| + + − − 3 | + + − − 14 | + + − − 18 |
| + − + + 12 | + − + + 3 | + − + + 3 |
| + − + − 23 | + − + − 5 | + − + − 0 |
| + − − + 10 | + − − + 35 | + − − + 35 |
| + − − − 22 | + − − − 97 | + − − − 102 |
| − + + + 46 | − + + + 2 | − + + + 4 |
| − + + − 40 | − + + − 13 | − + + − 14 |
| − + − + 64 | − + − + 26 | − + − + 24 |
| − + − − 58 | − + − − 111 | − + − − 110 |
| − − + + 160 | − − + + 5 | − − + + 2 |
| − − + − 188 | − − + − 38 | − − + − 9 |
| − − − + 1,004 | − − − + 28 | − − − + 31 |
| P<0.0323 | P<0.0593 | P<0.0184 |
Not: Çizim yeniden-yapılandırmalarında kullanılan bilgilendirici örüntüler kırmızıyla gösterilmiştir. Tek üyeli örüntüler yani (− + − −) ve (− − + −) gibi olanlar bu yeniden yapılandırmaları etkilemez. Kırmızı sayım kümelerinin istatistiksel olarak önemli olduğu olasılıklar, her bir sütunun altında verilmiştir.
Tablo.1’in sütunlarında gösterilen verileri yorumlamak amacıyla, ilk olarak, bilgilendirici sayımların en küçük kümesi ile o sütundaki ikinci en küçük bilgilendirici sayımlar kümesi arasındaki istatistiksel olarak en geniş aralığı belirledik. (Not: tek üyeli varlık/yokluk örüntüleri, yani + − − − veya − + − − gibiler, topolojik olarak bilgilendirici olmadığından analizlerimizde kullanmadık.) Soldaki sütunun yeniden-yapılandırımında, küçük sayımlar ile orta ölçekli sayımların kümesi arasındaki en büyük istatistiksel aralık 23 ile 40 arasında olup, CKO’nun sınırını tanımlar. Bu aralığın şans eseri olma olasılığı P<0,0323’tür. Dolayısıyla, bu sütunda 23 veya daha az olan sayımlarla desteklenen tüm tek üyeli örüntüler bilgilendirici olup, bu sayımlar rRNA’ların, mRNA’ların, tRNA’ların ve Zarların ortaklaşa evrimini tanımlayan halkaları yeniden yapılandırmak için kullanılmıştır.
Fig.3, Dünya’daki canlılığın kökeni ve evrimindeki başlıca dönemleri resmetmektedir. Bunlar, yatay çizgilerle bağlantılandırılan üç Süper Ortam olarak gösterilmiştir. Soldaki kırmızı çizgi CKO’yu SEOA’dan ayırırken, sağdaki pembe çizgi de SEOA’yı Canlılık Halkaları’ndan ayırır. Gen akışları zamanla arttığından CH’de büyük gen akışları vardır; SEOA’da orta ölçekli gen akışları vardır ve CKO’da küçük gen akışları vardır. Fig.3’te gösterildiği gibi, SEOA ile CKO’daki küçük gen akışları arasında istatistiksel olarak önemli aralıklar mevcuttur. Dolayısıyla, bu üç ortamın bilgisi, CKO’daki yaşamı yeniden yapılandırmamızı sağlar.

Canlılığın Kökeninin Yeniden Yapılandırılması
SEOA, CH’yi CKO’ya bağlantılandıran organizmalar topluluğudur. Fig.3’te CH ile CKO arasında bulunan daire ile temsil edilmiştir. SEOA’nın CH ile CKO arasındaki konumu, böylelikle CKO’yu Canlılık Halkaları’yla ilişkilendirmek için bir referans noktası sağlar.
CKO’daki evrimin yeniden yapılandırmak, CH’yi yeniden yapılandırmak için kullanılan tekniklerin bir modifikasyonuna dayanmaktadır (Lake et al. 2015). Bu yeni teknikler, bir sonraki bölümde açık bir şekilde tanımlanmıştır.
CKO Halkalarını Yeniden Yapılandırmak için Protein Ailesi Tanımlayıcıların Kullanılması
Canlılık CKO’dan SEOA’ya evrilirken Protein ailelerinin sayıları zamanla düzenli olarak arttığından, Fig.3’te gösterildiği gibi, biz ilk önce, Pfam örüntülerinin en küçük kümesini siyahla gösterilen daha büyük sayımlardan ayıran istatistiksel olarak önemli aralığı (Tablo.1’de kırmızıyla gösteriliyor) belirledik. Bu kırmızı, bilgilendirici sayımlar, Canlılığın Kökeni’nin halkalarının topolojisini benzersiz olarak tanımlar ve CKO’daki yaşamı yeniden yapılandırmak için kullanılmıştır.
CKO’nun Yeniden Yapılandırılmasındaki Adımlar
CKO’nun yeniden yapılandırılması dört adım içerir. İlk adımda, gen akışlarının en küçük kümesini belirliyoruz. Bu küme, daha büyük gen akışlarından, χ2 analizleri kullanılarak tanımlanabilecek olan istatistiksel olarak önemli bir aralıkla ayrılıyor. En küçük sayımların kümesi belirlenince, bu veriyi sanki SEOA’dan başlayarak modern Canlılık Halkaları’nı yeniden yapılandırıyormuşuz gibi analiz ediyoruz (Fig.4a). Ancak, en küçük gen sayımları, yani CKO sayımları yeniden yapılandırılmakta olduğundan, gen akışlarını zamanda tersine çevirmemiz de gerekiyor ki böylece genler CKO’dan SEOA’ya aksın. Bu, Fig.4a ile Fig.4b arasındaki geçişle gösterilmiştir. Üçüncü adımda ve son adımda ise CKO 180 derece döndürülerek, Fig.4c’de gösterilen geleneksel görüntü elde edilir. Bu zamanda tersine çevirmeli yeniden yapılandırma süreci kullanılarak, Canlılık Halkaları’nı yeniden yapılandırmak için kullanılan yöntemlerin sadece basit grafiksel modifikasyonlarını kullanarak, CKO’daki evrimi yeniden yapılandırabiliriz (Lake et al. 2015).

Bu işlem başarılıdır çünkü halka yeniden-yapılandırmaları, Pfam evriminin popülasyon davranışının dikte ettiği gözlemi temel almaktadır: a) evrimsel gen akışları içinde bulunan Pfam’ların sayısı zamanla sürekli artacaktır ki böylece Pfam’lar neredeyse hiçbir zaman Pfam gen akışlarından yitirilemez ve b) gen akışları o hâlde boyut açısından en küçükten (en eskiden) en büyüğe (en yeniye) düzenli olarak artar.
Geleneksel halka analizlerinde, SEOA ile CH arasındaki aralığı, Canlılık Halkaları’nın daha büyük sayımları ile SEOA’nın daha küçük sayımları arasında bulunan istatistiksel olarak en önemli Pfam aralığını bulmak için χ2 analizleri kullanarak tanımlıyoruz. Benzer biçimde, Canlılığın Kökeni araştırmalarımızda, CKO’nun daha küçük sayımları ile SEOA’nın daha büyük sayımları arasında bulunan istatistiksel açıdan önemli aralığı bulmak için χ2 analizleri kullanıyoruz. Daha sonra, CKO’daki evrimi yeniden yapılandırmak için küçük olan CKO sayımlarını kullanıyoruz.
Fig.4’te gösterilen yöntem, minimum filogenetik manipülasyon gerektirmektedir. İlk adımda, Fig.4a’da yeniden yapılandırılan halkada gösterilen okların hepsi ters çevrilir; böylece gen akışımız, Fig.2’de geliştirilen ve tanımlanan popülasyon biyolojisi modeline uygun olarak sürekli olarak artar. İkinci yeniden yapılandırma adımında, Fig.4b SEOA’nın merkezi etrafında 180 derece döndürülür; böylece Fig.4c’de gösterildiği üzere geleneksel görünüşte olduğu gibi CKO şeklin altına ve CH şeklin üstüne döndürülür. Dolayısıyla, halkaları döndürerek ve zamanın yönü tersine çevrilerek, CKO’nun evrimini yeniden yapılandırmak için geleneksel halka yöntemlerini kullanabiliriz.
Sonuçlar
RNA ve DNA Dünyalarının Yeniden Yapılandırılması RNA dünyası hipotezinden (Gilbert 1986) daha ilgi çekici ve tartışmalı olan hipotezlerin sayısı azdır; CKO’daki evrimi yeniden yapılandırabiliyor olmak ise bu kuramı sınamak için bir fırsat sunar. RNA dünyası kuramı şunu söyler: İlk nükleik asit organizmalarında DNA’dan önce RNA bulunmuş olabilir. Alternatif olarak, önce DNA bulunmuş olabilir ya da iki molekülün birlikte-evrimi olasıdır. Bu kuramları sınamak amacıyla biz, RNA’larda bulunan dört standart bazın (Adenin, Guanin, Urasil, Sitozin) evrimini tanımlayan halkaları yeniden yapılandırıyoruz ve bunları, DNA’larda bulunan dört standart bazın (Adenin, Guanin, Timin, Sitozin) evrimini tanımlayan halkalarla karşılaştırıyoruz.
Fig.5’te gösterildiği gibi, RNA-dünyası bazları ve DNA-dünyası bazlarının evrimsel yeniden-yapılandırımları aynı topolojiye sahip ve dolayısıyla her iki yeniden-yapılandırım da aynı halka kümelerini destekliyor. Tek fark şu ki RNA gen akışı bir Urasil ile başlarken, DNA gen akışı bir Timin ile başlıyor. Diğer tüm yönlerden, birleşik Timin/Urasil gen akışı oluşturacak şekilde iki akış birleşir birleşmez, ikisi de aynı evrimsel yolları izliyor.

Timin ve Urasil tarafından temsil edilen DNA ve RNA gen akışları birleşince, ortaya çıkan akışlar iki dala ayrılıp (bifürkasyon) Fig.5’te bulunan merkezi halkayı oluşturmaya başlamıştır. İlk önce Adenin yukarıdaki U/T akışına katılarak A/U/T gen akışı ve Sitozin aşağıdaki U/T akışına katılarak C/U/T gen akışı oluşturmuştur. Daha sonra, Fig.5’te sol altta gösterilen Guanin gen akışı iki dala ayrılarak, bir dal yukarı akmış ve A/U/T gen akışına katılmış ve nihayetinde SEOA’ya giden G/A/U/T gen akışını üretmiştir. Guanin gen akışının alt yolu T/U/C gen akışına katılmış ve bu iki akış birlikte G/T/U/C gen akışını oluşturmuştur. Daha sonra, G/T/U/C akışı A/T/U gen akışıyla birleşerek, G/T/U/C/A gen akışını oluşturmuştur (Fig.5’in sağ üst çeyreğinde gösterilmiştir). Bu akış nihayetinde figürün sol üst çeyreğinden gelen A/U/T akışına katılmış ve ardından bu iki gen akışı figürün üstündeki SEOA’yı oluşturmak üzere birleşmiştir. Bu sonuçlara ilişkin yorumumuz Tartışma Bölümü’nde belirtilmiştir.
Zarların, mRNA’ların, tRNA’ların ve rRNA’ların Evriminin Yeniden Yapılandırılması Zarların, mRNA’ların, tRNA’ların ve rRNA’ların kökenleri Fig.6’da yeniden yapılandırıldığı şekliyle gösterilmiştir. Bu halkalar, rRNA’nın halkaya giriş noktasından her iki yönde de akabileceği fakat mRNA, tRNA ve zar gen akışlarının giriş noktalarından sadece tek yönde ilerleyebileceği gözlemlenerek kurulmuştur. Ribozomal RNA (rRNA) gen akışı Fig.6’nın altına doğru ilerledikçe, zar gen akışıyla birleşir ve ilk rRNA içeren zarla çevrili hücreleri meydana getirir. Bu sonuçlar, ribozomların işlevsel yapı iskelelerinin kritik bir parçası olan rRNA’nın mRNA’lardan, tRNA’lardan ve zarlardan önce var olduğuna ilişkin güçlü kanıtlar (P < 0,0323) sağlar (Blattner et al. 1997; Simonson & Lake 2002; Tamura & Schimmel 2003; Ramakrishnan 2014).

Tartışma
İlk Genetik Kodlar Fig.5’te yeniden yapılandırılan RNA ve DNA gen akışlarından, en eski genetik kodların, bazların belirdiği zamansal sıralamaya dayandığı çıkarımını yapabiliriz. Bu gen akışları, en eski RNA/DNA genetik kodlarının ya UA/TA ya da UC/TC kodları olduğunu öngörmektedir ama UA/TA’nın mı yoksa UC/TC kodlarının mı önce evrildiğini belirleyemiyoruz. Sonraki kodların ilk olduklarını varsaymak sadelikli (İng. parsimonious) olmayacağından, biz en sadelikli kodların evrimini tanımlıyoruz.
RNA dünyası ve DNA dünyası halkaları içinde Urasil ile Timin’in önemi Fig.7’de görülmektedir. Urasil ile Timin kendi halkalarında ilk beliren bazlardır ve dolayısıyla RNA ve DNA dünyalarının başlangıçlarını tanımlarlar. Urasil, başka bazlarda olmayan çok sayıda dikkat çekici özelliğe sahiptir. Urasil’in geleneksel, titrek ve/veya başka sıra dışı baz çiftlenmeleri yoluyla Adenin, Guanin ve Sitozin ile baz çiftlenmesi yapma yetisi vardır (Tanaka et al. 2000). Dolayısıyla UA ve TA genetik kodları, ilk kodlar olmak için kabul edilebilir adaylar gibi görünmektedir.

Bu RNA bazlarının çift olabileceğini bildiğimizden, en eski gen akışlarında var olan bu nükleotidleri kullanan ilk genetik kodları sadeliklice öngörüyoruz. Spesifik olarak, ilk RNA genetik kodlarının aşağıdaki göreli beliriş sıralamasını gözümüzde canlandırıyoruz. Yeniden yapılandırılan RNA halkalarının içerdiği kronolojik bilgi, ilk iki genetik kodun UA ve UC kodları olduğunu sadeliklice öngörmektedir.
Fig.7’de gösterilen UA kodu, potansiyel olarak altı amino asidi kodlayabilirdi: fenilalanin, lösin, tirozin, asparagin, lizin ve izolösin. Bu altı amino asit şu seçkin kimyasal özelliklere sahiptir: Bunlar sırasıyla kutupsuz-hidrofobik, kutupsuz-hidrofobik, kutuplu nötr, hidrofobik nötr, asit hidrofilik ve bazik hidrofilik amino asitlerdir (Dickerson & Geis 1969) ve dolayısıyla bu “organizmalar” çeşitli ortamlarda potansiyel olarak işlev göstermiş olabilir. UC kodunun başlangıçta bir durdurma kodonundan yoksun olduğu öngörülmüş ve bu yüzden protein sentezinin sonlandırılmasının verimsiz olacağı varsayılmıştır. Ancak, UC akışı daha sonraları UA gen akışıyla birleşince bir durdurma kodonu kazanmış olabilir. Ek olarak, UA kodu UAA durdurma kodonunu içerir (turuncu). Bu durdurma kodonu, UA organizmalarının belirli bölgelerde protein sentezini sonlandırmasını ve çerçeve kayması mutasyonlarını baskılamasını olanaklı kılmış olabilir. Daha sonra, GUG ve UUG gibi başka durdurma kodonları da belirmiştir ama bizim filogenetik yeniden yapılandırmalarımız sadeliklice işaret ediyor ki UAA durdurma kodonu çok eskiydi ve muhtemelen ilkti. Eğer öyleyse, bu UA soyundaki kritik bir yenilik anlamına gelir.
UC kodunun dört amino asidi kodladığı sadeliklice öngörülmektedir: fenilalanin, lösin, serin ve prolin; çünkü genetik kodda herhangi bir amino asidin başka bir amino asitle yer değiştirdiğine ilişkin hiçbir kanıt yoktur. Kimyasal olarak, bu amino asitler sırasıyla kutupsuz-hidrofobik, kutupsuz-hidrofobik, kutuplu nötr ve kutupsuz-hidrofobiktir.
UA kodu gibi, UC kodu da bir AUG başlatma kodonu içermez. Dolayısıyla, bu çok eski organizmaların, şu anda protein sentezini başlatmak için kullanılan modern genetik mekanizmadan yoksun olduklarını öngörmekteyiz. Bununla birlikte, grafiğin sol üst tarafından SEOA’ya giren UAG gen akışı gibi veya grafiğin sağ üst tarafından SEOA’ya giren UACG gen akışı gibi daha sonraki kodlar, AUG başlatma kodonları için potansiyel kaynaklardır. Dolayısıyla, bu sadeliklice öngörür ki, protein sentezinin başlatılmasının kontrolü, CKO sırasında canlılığın evriminde nispeten geç bir zamanda belirmiştir.
İlk Hücrelerin Kökeni: rRNA Dünyasının Zar Dünyasından Eski Oluşu İlk zarın kökeni, canlılığın evriminde çok önemli bir adımdı. Bu evrimsel ilerleme, hücreseli hücresel olmayan yaşamdan ayırdı. Zarlar, hücrelerin içindekileri seyrelmekten, değiştirilmekten ya da dış çevre tarafından başka olumsuz şekillerde etkilenmekten korumayı olanaklı kıldı. Biyokimyasal canlılığın kökeni deneyleri göstermiştir ki çok sayıda dışsal ısıtma ve soğutma çevrimleri yoluyla basit zar veziküller içinde DNA’yı modifiye etme olanağı vardır; böylelikle “…veziküller içinde tutulurken, çift-iplikli DNA yüksek sıcaklıklarda denaturasyonla ayrılabilir (Mansy & Szostak 2008),” ve benzer biçimde “RNA, protein ve lipid prekürsörlerin” ortak kökenleri gösterilmiştir (Patel et al. 2015). Zarlar ilk hücreyi kaplar kaplamaz, beliren canlılık sitoplazmik ortamını düzenlemeye başlayabilmiştir.
Ancak şimdiye dek, zarların mı yoksa RNA’ların mı önce geldiğine ilişkin herhangi bir filogenetik kanıt olmamıştı. CKO’daki genomiğe dayalı filogeniler üretmemizi sağlayan yeni becerimiz düşünülürse, artık RNA’ların zarlardan önce mi yoksa sonra mı geldiğini sorabiliriz. Eğer RNA’lar DNA’dan önce geldiyse, hangi RNA’ların (mRNA, tRNA ya da rRNA) ilk olduğunu da bilmek isteriz.
İlk Zarla Çevrili Hücreler
Canlılığın kökeni deneyleri, ilk zarların önemini açıkça göstermiştir ve sağladıkları büyük avantajları açığa çıkarmıştır (Mansy & Szostak 2008). Bu da zarların edinilmesinin, bu ilk hücrelerin belirmesi için güçlü bir seçilimsel avantaj ürettiğini akla getirmektedir. Ancak şimdiye dek, ilk hücrelere götüren evrimsel adımlara ilişkin doğrudan filogenetik kanıtın olmaması, rRNA’lar, mRNA’lar, tRNA’lar ve zarlardan hangisinin ilk önce geldiğini belirlemeyi olanaksızlaştırmıştır. CKO’nun filogenetik yeniden-yapılandırmalarını kullanarak, artık bunlara ve benzer sorulara doğrudan eğilebiliriz. Şu anda, rRNA’nın mRNA, tRNA ve zarlardan daha eski olduğuna ilişkin istatistiksel olarak önemli kanıta sahibiz.
Sinerjik Canlılığın Kökeni Araştırmalarının Geleceği Canlılığın kökenindeki başlıca olayların yeniden yapılandırılmasının geleceği parlak görünmektedir çünkü Fig.8’de gösterildiği gibi Dünya’daki evrimin birleşik bir görüntüsünü sağlar ve ayrıca biyokimyasal süreç kanıtlama deneyleri artık CKO’daki filogenetik halka yeniden yapılandırmalarıyla doğrudan karşılaştırılabilir. Canlılığın kökenini yeniden yapılandırmak ve anlamak için kullanılan bu yeni filogenetik aracın yeni keşiflere kapı açabileceğini öngörüyoruz. Gelecekteki ilerlemelerin, deneysel canlılığın kökeni araştırmalarında uzman olan laboratuarlar ile CKO’daki genom evriminin yeniden yapılandırılmasında uzman olan laboratuarlar arasında sinerjik ortaklıklar içereceğini umuyoruz.

Böyle ortaklıkların, nükleik asitlerin, protein sentezinin, zarların ve amino asitlerin evrimini kavrayışımızı bütünleştirme potansiyeli bulunmaktadır; biz onların canlılığın kökeninin yeni evrimsel sentezini üretebilme potansiyeli olduğunu ileri sürüyoruz. CKO’nun ileride yapılacak yeniden yapılandırmalarının yeni canlılığın kökeni yollarını açığa çıkarmasını ve Dünya’daki canlılığın evrimi sırasında gerçekleşen biyokimyasal, enerjik, yerbilimsel ve genomik olayların daha kapsamlı, bütünleşik bilgisini sağlamasını bekliyoruz.
Geçmişte, canlılığın kökenini yeniden yapılandırmak, filogenetik çözümlemelerin eriminin ötesinde gibi görünüyordu ve SEOA’dan önceki evrimi yeniden yapılandırmanın olanaksız olduğu evrensel olarak varsayılıyordu. Başlangıçta, amino asitlerin abiyotik sentezi (Miller 1953) ve RNA dünyası hipotezi (Gilbert 1986) gibi başlıca ilerlemeler, CKO’daki erken dönem olaylarını günün birinde anlayabileceğimize ilişkin umut vermişti. Zaman içinde, amino asitlerin (Miller 1953), zarların (Mansy & Szostak 2008) ve RNA, protein ve lipid prekürsörlerin (Patel et al. 2015) oluşumu da dahil olmak üzere, canlılığın kökenindeki erken dönem olaylarını açıklamaya yardım eden uygulanabilir biyosentetik yolaklar keşfedildi. Burada tanımlan yeni genomik yöntemlerin bu çalışmaları tamamlamasını, yeni araştırma doğrultuları göstermesini ve ilave başlıca biyolojik canlılığın kökeni geçişlerini kronolojik olarak sıralamamıza olanak tanımasını bekliyoruz. Genomik, biyokimyasal-uygulanabilirlik deneyleri, erken dönem Dünya’nın paleontolojik araştırmaları ve başka gezegenlerdeki araştırmaların sinerjik birlikteliklerinin, CKO’daki canlılığın evriminin keşfi ve kavranmasındaki ilerlemeleri önemli ölçüde ivmelendireceğini öngörüyoruz.- Uncovering the Genomic Origins of Life - J.A. Lake & J. Larsen & D.T. Tran & J.S. Sinsheimer (Genome Biology and Evolution - 2018 Jul; 10(7): 1705–1714.) https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6047450/
Dilediğiniz miktarda aylık veya tek seferlik bağış yapabilirsiniz.
Destek OlBunlar da İlginizi Çekebilir






En Çok Okunan
Bu Ay Öne Çıkanlar



İnsanlık Uygarlığı Neden Bu Kadar Geç Keşfetti?

E-Bülten Üyeliği
Duyurulardan e-posta ile haberdar olmak istiyorum.